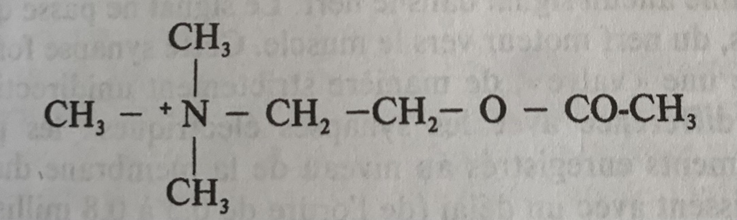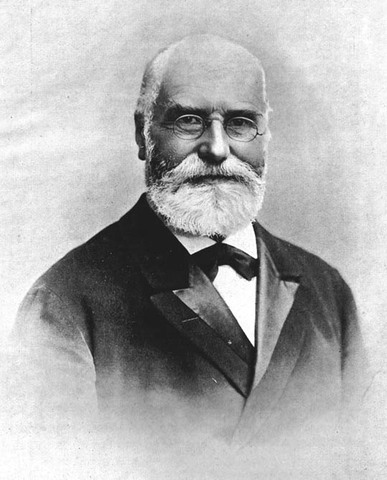Pour introduire la lecture de ce livre assez récent paru en 2023 aux éditions Seuil, je vais vous faire part d’une petite anecdote la dernière fois que je suis sorti dans Paris.
Je me rappelle avoir interpellé une jeune fille qui avait l’air perdue. Elle me dit : « je cherche mon bus pour me rendre dans le 91 ».
On était la nuit vers 23h30 et en me montrant les panneaux « Noctilien » tout en consultant son téléphone mobile et un service de géolocalisé de type Google ou Plans. Elle ne comprend pas son bus n’est pas à l’endroit ou il devrait être. Je lui tend le bras vers l’arrêt: ça c’est la N, elle me rétorque « Nuit, Noctilien » je rigole et je reprend : « L’arrêt en face de toi c’est le réel » ensuite je lui désigne son téléphone et je reprend de plus belle, ça c’est « virtuel ».
Elle fait demi-tour et elle me dit : « tu as raison ».
Je trouvais l’anecdote amusante pour vous introduire ce livre de Jean-Gabriel Ganascia dont on avait déjà parlé dans mon article sur les Sciences Cognitiveset qui fait montre de toute son éminence grise dans ce livre intitulé Servitude Virtuelle. Quel titre ! Tout un programme n’est-ce pas ?
Et pourquoi pas ne pas commencer par la fin du livre et faire une analogieavec le discours de la servitude volontaire d’Etienne de la Boétie écrit 400 années plus tôt ?
« Ils s’en servent par forme et pour épouvantail plus qu’ils ne s’y fient. Les archers barrent l’entrée des palais aux malhabiles qui n’ont aucun moyen de nuire, non aux audacieux bien armés. Ce sont les peuples eux-mêmes qui se laissent, ou plutôt qui se font malmener , puisqu’ils en seraient quittes en cessant de servir. C’est le peuple qui s’asservit et qui se coupe la gorge ; qui pouvant choisir d’être soumis ou d’être libre, repousse la liberté et prend le joug ; qui consent à son mal, ou plutôt le recherche… »
Le qualificatif de « virtuel » qui nous intéresse pour cette nouvelle forme de servitude : se réfère aux sciences du numérique et vient s’opposer au réel. Sommes nous aujourd’hui perdus entre deux mondes ? La clé réside t-elle dans l’hybridation ?
Doit-on se séparer de nos téléphones mobiles quand on se déplace de peur d’être enfermé dans une toile virtuelle, (ondes électromagnétiques (radio) captées par les antennes du réseau cellulaire) (Puces de géolocalisation par satellite) . Doit-on pour s’orienter ne se fier qu’à nos connaissances astrales (Soleil, Etoiles, Cartes, Points Cardinaux..) ? Doit-on faire confiance aux outils d’intelligence artificielles ? Aux algorithmes de recommandations ? A la diffusion Point par Point ? Au Marketing one-to-one qui pardonnez moi ce néologisme nous « enbulle » ?
Ce site scientifique et pédagogique que j’ai crée vous donne toutes les clés pour comprendre le numérique, les sciences qui en retournent et ce livre « Servitudes Virtuelles » rentre dans cette démarche . J’ai voulu ainsi vous en dévoiler une partie de son contenu.
L’auteur tente de répondre à ces questions en abordant des thèmes très actuels et démarre par un fabuleux concept :
1ère Partie :
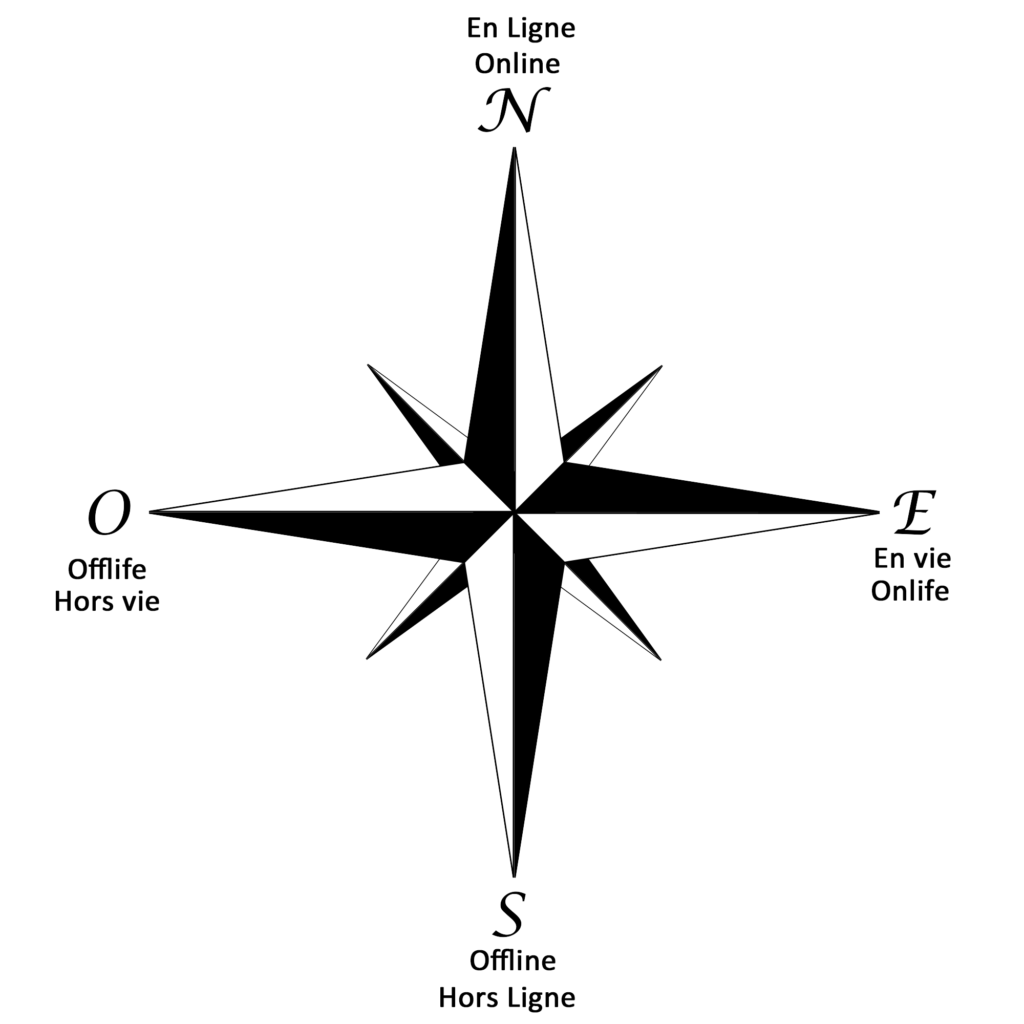
Une rose des vents numérique :
Dans la nature, la rose des vents était censé guider les marins pour la navigation à l’aide de ses pétales. Encore une preuve que la nature a tout inventé et la technologie ne fait que la copier comme je l’ai déjà montré dans mon article sur l’appareil cérébral.
Pourtant nous vivons dans une époque ou les mathématiques rêvent de contrôler la nature.
1/Hors-ligne, Off-life, l’ouest. (Là ou le soleil se couche)
L’auteur commence à l’ouest, cet état est dis Off-life, Hors-life il y parle de dataïsme concept inventé par le chercheur homosexuel israëlien Harari, et repris par Chris Anderson du célèbre magazine Wired qui pense que la théorie va disparaître en conséquence.
Ensuite l’auteur parle de métensomatosenumérique c’est à dire de transfert de l’esprit dans le corps humain par téléchargement pour au final arriver à l’état de mort décidé à par un agent artificiel, un programme ou un ordinateur. L’auteur plagie volontairement 4 passages d’un roman transhumaniste dont vous l’aurez compris nous ne sommes finalement pas loin aujourd’hui en 2025.
2/ Online, le Nord. (L’étoile Polaire, Vénus)
Ensuite dans un deuxième temps nous sommes dans le Online, en ligne. La vie numérique par excellence.
Dans l’online l’auteur nous apprend que l’ancêtre du protocole hypertexte date de la seconde guerre mondiale en 1945 par un physicien du nom de Vandevar Bush qui a écrit un traité intitulé As We May Think ?
L’auteur y parle des projets Neuralink d’Elon Musk nommé par le syntagme dentelle neurale.Pour résumer tous ensemble nous ne formerons plus qu’une fratrie connecté par une puce qui nous augmentera et nous permettra de contrôler l’Intelligence Artificielle.
Mark Zuckerberg souhaite aussi mieux connaître ce que les personnes pensent pour mieux profiler et mieux personnaliser la publicité.
Ce sont des projets dont les neuropsychologues doutent de la faisabilité pour la raison qu’il y’a 3 phases pour accéder à la mémoire : encodage, consolidation et accès.
3/ En vie, Onlife, l’Est. ( Là ou le soleil se lève)
L’auteur fait mention de la blockchain, idéal cyberpunk de hack, mathématisation binaire de l’argent en blocs cryptographiques et diverses fonctions de hachages censés renverser l’ordre établi et se passer du système bancaire. Rappelons que la blockchain a été inventé en 2008 après la crise des subprimes par un japonais répondant au pseudonyme de Nakamoto.
Est évoqué Hannah Arendt grande philosophe juive expatrié aux Etats-Unis dont sont mentionnés les travaux sur la culture, la vie contemplative, la vie active, la vie de l’esprit (Cultura Anima).
Sont évoqués la culture de masse, la reproduction mécanique des œuvres, leur transports et leurs diffusion. Mais aussi le travail, l’œuvre et l’action pour un homme.
Le taylorisme a l’extrême avec le morcellement du travail de plus en plus grand et l’exemple des Amazon Mechanical Turks. Ce sont de petites tâches très peu rémunérés qui étaient faites par de petites mains qui auraient très bien pu être remplacés par des IA codées pour gagner de l’argent facile. Max Tegmark (Life 3.0).
4/ Enfin le Offline ou le Hors Ligne. Le Sud.
L’auteur évoque la ZAD de notre dame des landes et des personnes complètement déconnectés du monde actuel mais qui se battent pour des causes tout à fait nobles et qui sont obligés finalement pour se faire entendre de faire une incursion dans le online.
2ème Partie :
Ensuite l’auteur nous invite à prendre un marteau comme vous pouvez le voir en couverture pour déconstruire tout cet univers numérique et il nous invite à réfléchir à 4 principes d’équité pour tenter d’orienter le monde dans la bonne direction.
1/Autonomie,
2/Bienfaisance et non Malfaisance
3/Justice
4/Transparence
Le livre se termine enfin en apothéose avec un petit trésor que je ne divulguerai pas pour vous forcer à l’acheter.
De plus il est très documenté et il ne faut absolument pas négliger les notes et leurs renvois lors de la lecture. Beaucoup de sources numériques de haut vol et universitaires sont inscrites dans l’index finale et valent le coup d’être explorées après ou pendant lecture.
Je vous recommande vivement l’achat de ce livre très actuel.
Ceci est un résumé du chapitre 3 de Changeux « L’homme neuronal » :
Accrochez-vous c’est à mon sens un des chapitres les plus denses du livre l’homme neuronal de Jean Pierre Changeux en termes de concepts mais il est fondateur !
Electroencéphalographie
Après avoir étudié les différentes parties du cerveau, la manière dont elles se structurent entre elles de manière typologique et topologique sur un plan statique. Passons à l’étude dynamique de leurs fonctions : comment la machine cérébrale organise la transmission de signaux à travers les différents organes sensoriels et moteurs.
Nous savons notamment que le cerveau fonctionne grâce à des impulsions électriques, commençons par une étude d’électroencéphalographie :
Hans Berger 1873-1941
Hans Berger plaça pour la première fois des électrodes sur le cerveau humain contrairement à Caton et Matteucci qui avaient pris pour cobaye des animaux. Ces électrodes ne pouvant être implantés directement dans le cerveau, elles sont apposées à la surface du crâne, la traduction du signal étant optimisé par un amplificateur de gain.
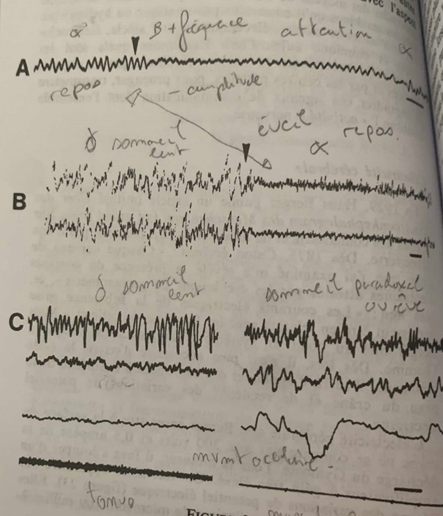
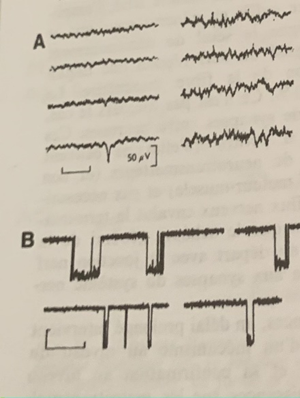
En A, d’abord les ondes alpha (repos) puis augmentation de la fréquence à partir de la flèche pour un passage aux ondes Beta : l’attention se fixe et retour aux ondes alpha. (Repos) En B, le sommeil lent et les ondes delta à forte amplitude et fréquence très lente, la flèche indique le passage à l’état de réveil avec un retour des ondes alpha (repos) En C, à gauche le sommeil lent avec les ondes delta, à droite le sommeil paradoxal (rêve) Le deuxième tracé indique le passage des ondes delta à un rythme paradoxal en rafale. Le troisième tracé lorsque les yeux bougent pendant le sommeil. Le quatrième le tonus musculaire qui s’affaisse lors du sommeil paradoxale (à droite)
A l’état de repos la fréquence des ondes électriques est lente et de 10 cycles par secondes et leur amplitude est forte. Ce sont les ondes alpha α. Au contact d’un corps solide telle une tige de verre, la fréquence des ondes devient plus forte, la fréquence est plus forte et l’amplitude diminue : l’attention se fixe, ce sont les ondes Beta β. #ondes
Lors du sommeil lent l’amplitude des ondes est encore plus forte et la fréquence encore plus faible qu’à l’état de repos. Ce sont les ondes delta δ. Lors du réveil on passe des ondes δ à α donc à l’état de repos. #ondes
Un autre état est celui de sommeil paradoxal ou rêve, contrairement au sommeil lent les impulsions électriques sont souvent en rafales. Les yeux bougent et le tonus musculaire s’affaisse. #ondes
Cependant cette étude s’avère beaucoup trop généraliste de par l’irrégularité des ondes β et la similitude de fréquence et d’amplitude des ondes δ et α pour comprendre les phénomènes intrinsèques qui se produisent au niveau cellulaire et moléculaire. #ondes
On doit alors passer au cerveau de l’animal pour pénétrer le cortex avec l’électrode, on obtiendra en une fois ce qu’on a obtenu avec les électrodes apposées sur le crâne humain dont les résultats furent cumulés. #ondes
En utilisant une électrode dont la pointe est entre 0,5 et 5 micromètres plus petite que celle de la cellule nerveuse et on obtient des impulsions très brèves dont l’intensité s’accroît puis s’évanouit (discrètes) lorsque l’on déplace la microélectrode sur toute la surface corticale.
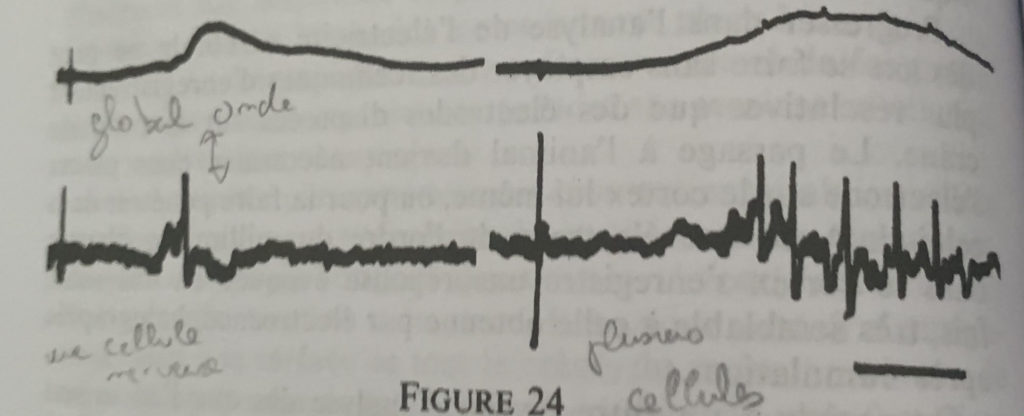
On répète l’expérience en plaçant l’électrode dans le cortex somatosensoriel, on stimule alors la patte du rat et l’on obtient une activité qui permet de mettre en lumière une source électrique forte continue et produite par des entités discontinues j’ai nommé le neurone.
En haut une one globale avec une legere polarisation, en haut a droite une polarisation plus forte mais toujours globale. La correspondance en bas permet de mettre en evideidence la microstructure du neurone.
Cette démarche encéphalographique permet donc la mise en relation de l’activité électrique avec sa microstructure (neurone) mais ne pourra rendre compte du fonctionnement électrique du neurone de par sa multitude et des interactions complexes entre chacun.
L’influx nerveux
La vitesse de propagation électrique de l’influx nerveux chez les animaux primitifs se déplacent de 0,1 m/s (méduses) contre 100 m/s pour certains mammifères ce qui reste inférieur à la vitesse du son mais peuvent atteindre le mètre. La membrane cellulaire du neurone conduit le courant et remplace les fils de cuivre dans une communication électronique.
La durée de l’impulsion ne varie pas, une période réfractaire intervient pour maintenir sa durée ainsi que pour espacer deux influx successifs. L’amplitude des impulsions d’un dixième de Volt ne varie pas non plus. Quel que soit l’endroit où on l’enregistre ou la manière dont il est produit l’influx nerveux à la même forme.
La communication dans le système nerveux se traduit par un système nerveux très uniforme d’impulsions électriques voir universel.
Schéma représentant le signal électrique d’une onde d’influx nerveux dans l’axone géant du calmar à gauche (directement dans l’animal) (à droite après dissection) On peut même expulser le cytoplasme de l’axone, le placer dans une solution saline et on obtient à peu près le même potentiel.
Si l’on pénètre maintenant avec la microélectrode dans le neurone même on obtient un potentiel électrique stable dit de repos. La valeur varie par rapport à celle de la périphérie (Axone) de 0,50 à 0,90 mV.
Comment expliquer cette différence de potentiel avec l’expérience précédente à la périphérie au niveau de l’axone dans lequel le potentiel électrique était plus fort ?
Le neurone est une pile électrochimique
Le neurone est constitué d’atomes de sodium et de potassium, cependant on relève dix fois moins de sodium à l’intérieur qu’à l’extérieur mais dix fois plus de potassium. La chimie se transforme en électricité par ionisation, la libération d’un électron (particule charge négative) de valence va transformer le sodium et le potassium en ions positifs Na+ et K+. La membrane va filtrer ces ions et ne laisser passer à l’état de repos que les ions potassium de part et d’autre provoquant une force électromotrice négative à l’intérieur.
L’énergie qui maintient les concentrations d’ions potassium et sodium est une molécule biochimique : l’adénosine triphosphate ATP produite par respiration cellulaire (phénomène par lequel la mitochondrie (métabolite) transforme par phosphorylation (phosphore) oxydative l’Adénosine Di(2)phosphate en Adénosine Tri(3)phosphate). L’ATP Synthase qualifiée d’enzyme pompe de l’ATP (le suffixe ase désigne toutes les enzymes) sectionne ensuite l’ATP et cette scission va servir d’énergie pour provoquer l’influx nerveux de part et d’autre de la membrane.
Au repos la membrane est donc quand même mise sous tension par l’ATP mais elle ne laisse pas passer les ions sodium. La théorie du repos a été défendue par Julius Bernstein
Julius Bernstein 1839-1917
et confirmée plus tard par Charles Overton qui théorisera l’effondrement de la membrane par scission d’ATP pour expliquer la génération d’impulsion nerveuse.
Charles Overton 1865-1933
Huxley et Hodgkin vont préciser de manière universelle tout ce que nous venons de voir (mécanique ionique de l’influx nerveux) à partir d’une série d’expérience en utilisant encore l’axone géant du Calmar. En effet la perméabilisation de la membrane aux ions sodiums est commandée par une valeur seuil de potentiel électrique.
Les ions sodiums s’engouffrent dans des molécules canaux et le potentiel passe d’un signe négatif à un signe positif + 20 mv. L’impulsion électrique générée dépasse l’amplitude de 100 mV le long de l’axone. Ces canaux vont se refermer, d’autres canaux pour le transport d’ions potassium vont s’ouvrir et l’on retournera à l’état de repos, cette impulsion électrique aura duré 1ms.
Huxley 1917-2012
Hodgkin 1914-1988
Ceci s’applique universellement du nerf sciatique du rat à l’axone du Calmar et au cerveau humain.
Les oscillations
On a vu que même à l’état de sommeil lent au niveau encéphalographie, l’activité électrique était présente même sans stimulation sensorielle. Les neurones mis en cultures et tumoraux ont quand même un potentiel électrique régulier. Ces oscillations témoignent d’une activité spontanée du neurone.
Les expériences réalisées sur l’aplysie, la limace de mer qui présente l’avantage d’avoir des neurones géants vont faire état d’oscillations en rafales d’une parfaite régularité d’une dizaine d’impulsions toutes les 5 à 10 secondes.
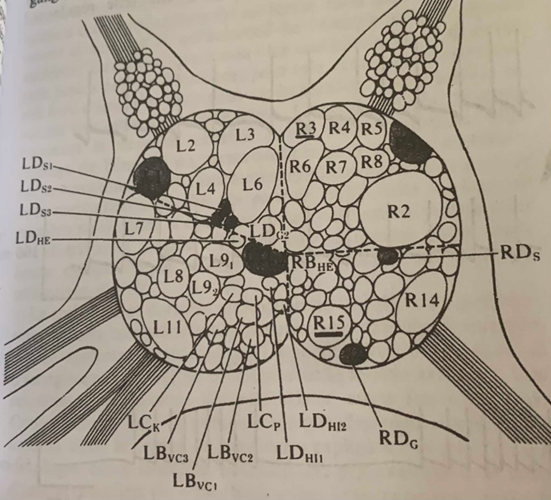
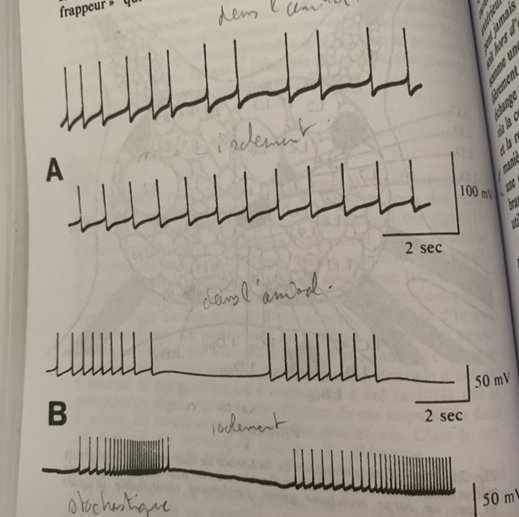
Le ganglion abdominal de l’Aplysie avec ses neurones géants, on étudie le R3 et le R15.En A l’horloge à rafale électrique spontanée du neurone R3, En B l’horloge à rafales régulières d’impulsions (stochastique) La ligne supérieur représente le schéma de l’implantation de l’électrode au niveau du neurone en gardant l’animal dans son entier. En dessous l’implantation de l’électrode dans le neurone isolé du reste de l’animal.
Ilya Prigogine s’empare de la question pour voir si ces oscillations cadrent avec les lois de la thermodynamique son domaine scientifique de prédilection. Voici les trois lois établies avant lui par Lord Kelvin à laquelle obéit cette science macroscopique :
Ilya Prigogine 1917-2003
1/ L’énergie est égale dans l’univers, on ne peut ni créer ni détruire, on peut faire l’analogie avec Lavoisier qui dit que rien ne se perd mais tout se transforme. Ainsi on peut résumer cette loi par le syntagme d’Echange universel.
2/ L’Entropie augmente toujours dans un système isolé, l’entropie se caractérise par l’énergie thermique qui ne peut être utilisé d’aucune manière. Au sens général l’entropie est une grandeur propre au désordre de la matière d’un système.
3/ La troisième introduit le zéro absolu -276,15 C° et dit qu’à cet état l’entropie est nulle. Mais on sait qu’aucun être vivant rien ne peut vivre à cette température cependant on comprend que plus on tend vers une entropie faible, plus l’état énergétique est stable.
On parle même d’état métastable un peu comme la supraconduction qui expulse le champ magnétique et fait flotter le train hyperloop au-dessus des rails.
Comment les oscillations du neurone cadrent-t-elle avec ses trois lois ?
Je mettrai des chiffres à côté des mots qui vous permettent de relier ces conclusions aux 3 lois.
Première conclusion : les oscillations du neurone appartiennent à un système ouvert (2) dans lequel il y’a échange (1) constant (3) d’énergie. (De par la perméabilité de la membrane, dans lequel l’énergie s’échange grâce à l’ATP)
Deuxième conclusion : Les oscillations ne se développent jamais près de l’équilibre, pour cela il faut que le système soit hors d’équilibre*mais stable (3). Le neurone répond parfaitement à ces deux conditions (transfert inégale et continu d’ions)
Troisième conclusion : Des relations non linéaires* (propre des oscillations) doivent exister entre forces et flux, concrètement cela se présente lorsque des réactions se développent de manière explosive et lorsque des couplages s’établissent entre entrées et rétroaction (feedback en cybernétique) Le déclenchement de l’influx nerveux satisfait à ces conditions.
*Pour illustrer ces conclusions il est bon de souligner que Prigogine s’est écarté à la fin de sa vie de la thermodynamique classique a établi le concept de structure dissipative : « une nouvelle ‘structure’ est toujours la conséquence d’une instabilité. Elle est engendrée par une fluctuation. »
La particularité de cette oscillation est sa rafale :
Chaque rafale se greffe sur un générateur d’oscillation(pacemaker, rythme) dont le potentiel électrique oscille entre deux valeurs extrêmes de part et d’autre du seuil d’apparition de l’influx nerveux. Lorsque la valeur seuil du potentiel électrique est franchie une impulsion part et puis une autre tant que le potentiel reste au-dessus de la valeur seuil.
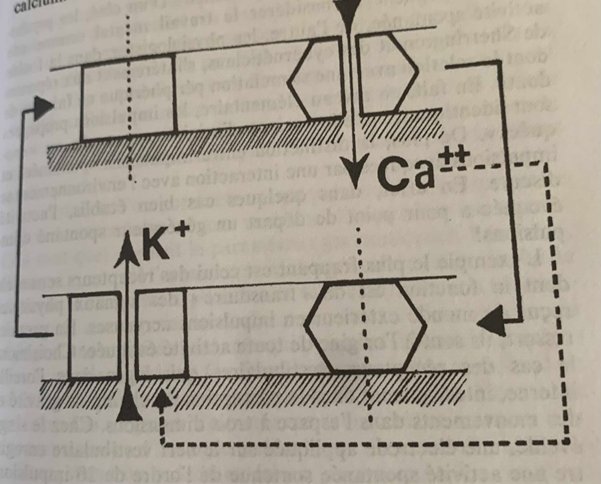
Comme pour l’influx nerveux on retrouve deux molécules canaux mais leur ouverture est de l’ordre d’une seconde pleine contre une milliseconde pour ce dernier. D’autre part le filtrage de la membrane du neurone est différent, les molécules-canaux filtrent les ions potassium et calcium (exclu de l’intérieur du neurone par une enzyme comme l’ion sodium) :
Schéma très simplifié de la boucle qui s’opère lors des oscillations de base générés par l’ouverture des molécules canaux Calcium et Potassium. Le calcium passe, est rejeté par l’enzyme pompe, le canal se ferme mais l’ion provoque l’ouverture du canal potassium, l’ion potassium passe et même le même schéma se répète.
La canal calcium très lent s’ouvre par diminution du potentiel électrique crée par le canal potassium et laisse passer l’ion Ca++ qui une fois rejeté par l’enzyme pompe va provoquer l’ouverture du canal potassium dont on récupère l’ion K+ et on recommence comme dans un cycle. Ce couplage rétroactif est donc en parfait accord avec la troisième conclusion de Prigogine.
Si l’oscillation a une amplitude suffisante pour que la valeur seuil de l’influx nerveux soit franchie une rafale part à la crête de chaque oscillation lente : la longueur de la rafale dépend de l’amplitude et de la durée de l’oscillation.
Si le temps est bref on peut n’avoir qu’une impulsion voir pas du tout.
On a donc soit une horloge à rafale soit un générateur stochastique d’impulsion déterminé par la concentration de calcium dans la membrane.
Les sens et l’activité spontanée
Les récepteurs sensoriels ont pour but de transduire les signaux perçus par le monde extérieur en impulsion électrique : Lorsque l’on fait tourner un singe assis sur une chaise auquel on a placé une électrode dans le nerf vestibulaire de l’oreille dans un sens : on obtient 30 impulsions. Dans l’autre sens on obtient plus que 10 impulsions.
L’organe vestibulaire se compose de neurones tournés en direction du système nerveux central et de cils palpeurs qui baignent dans du liquide. Lorsque l’on tourne d’un côté la membrane va être orienté de telle façon que les canaux calciums s’ouvrent : diminution, ou se ferment : accroissement.
La variation d’un paramètre physique (gravitationnel, chimique ou lumineux) se trouve donc traduite en variations d’impulsions nerveuses. Les organes des sens se produisent comme des commutateurs d’horloge moléculaire. Les stimuli physiques les avancent, les mettent à l’heure où les retardent.
Les propriétés de générateurs d’impulsions ne sont pas réservées aux cellules sensorielles Il s’agit d’une propriété générale de la cellule nerveuse distribuée à de multiples niveaux.
La machine nerveuse contient donc de multiples générateurs d’impulsions distribuées tant au niveau du système nerveux lui-même que dans son ensemble.
D’un neurone à l’autre
Le sens de propagation des impulsions électriques est unidirectionnel, il part du corps cellulaire jusqu’à la terminaison de l’axone puis les dendrites comme dans un circuit électronique avec des sens obligatoires et des sens interdits.
On sait que les neurones et dendrites ne sont pas toujours en continuité les uns avec les autres. Certains neurones sont juxtaposés et séparés les uns aux autres. Comment peut-on expliquer la transmission du courant dans cette organisation discontinue ?
Comment expliquer cette polarité étant donné que lorsque l’on induit du courant électrique au milieu de l’axone le courant part dans les deux sens ?
Sherrington dont on a parlé dans l’histoire des neurosciences l’explique par la synapse en comparant un tronc nerveux ou le signal est bidirectionnel et l’arc réflexe unidirectionnel (stimuli sensorielle entraînant une réaction nerveuse comme un mouvement). Pour lui la synapse est une barrière intercellulaire : un nexus (espace proche) entre deux neurones.
Sherrington 1857-1952
Ceci va engendrer un débat entre les pharmacologues pour lesquels la transmission est chimique et les électro-physiologistes qui soutiennent la thèse électrique. Un compromis est trouvé : il existe des synapses électriques et des synapses chimiques.
Les synapses électriques sont symétriques et se placent entre deux neurones contigus de 2 nanomètres d’épaisseur, il y’a communication directe entre les neurones la plupart du temps sans polarité de manière multidirectionnelle comme une horloge qui mettrait les neurones à la même heure.
La transmission chimique est paradoxalement découverte par un électro-physiologiste dont on a déjà parlé dans l’article premier Du Bois Reymond mais aussi aux pharmacologues Elliott, Sir Henry Dale, et Langley qui prélèvent sur certains organes cibles de la grenouille un ester de la choline, l’acétylcholine.
Thomas Renton Elliott 1877-1961
Sir Henry Dale 1875-1968
Emil Heinrich Du Bois Raymond 1818-1896
Ce neurotransmetteur a été découvert dans la nature à partir de l’ergot de seigle et synthétisé par Crum-Brown et Frazer
Alexander Crum Brown 1838-1922
James Frazer 1854-1941
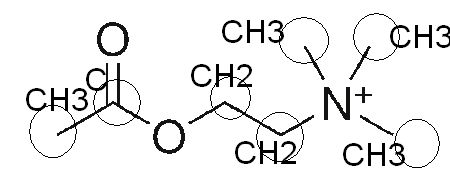
Faisons un petit exercice de biochimie en considérant la formule brute, la formule semi-développée et la formule développée de l’acétylcholine en partant de la composition d’un ester et de la choline.
Formule Brute Acétylcholine : C7,H16,N+O2
On trouve donc 7 atomes de carbone, 16 atomes d’hydrogènes, un atome d’azote, et deux atomes d’oxygène.
Voici la formule semi-développée qui présente une première disposition dans l’espace de manière rectiligne ainsi que les liaisons entre les atomes.
Formule semi développée de l’acétylcholine (Basé sur l’ion positif d’Azote de symbole atomique N)
Comparons avec la choline formule brute C5,H14,N+O :
On retrouve bien le nombre d’atomes énoncés dans la formule brute
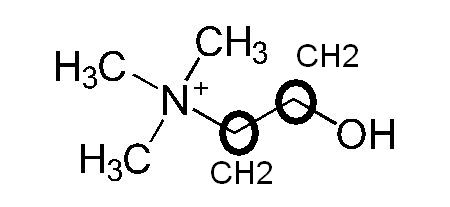
J’ai complété la formule développée en deux dimensions que l’on trouve sur wikipedia à partir de l’énoncé de la formule brute avec des cercles et des liaisons atomiques.
on retrouve les liaisons CH3 autour de l’atome d’azote, les deux sommets cerclés n’étaient pas notés atomiquement, je les ai rajouté.
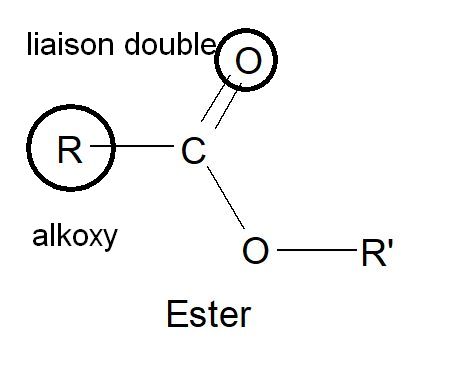
En chimie, la fonction esterdésigne un groupement caractéristique formé d’un atome lié simultanément à un atome d’oxygène par une double liaison et à un groupement alkoxy
liaison double deux traits lie l’atome de carbone à l’atome d’oxygène, alkoxy est symbolisé par le groupement R et R’
Lorsque l’atome lié est un atome de carbone , on parle d’ester carboxylique, dont la forme générale est R-COO-R’.
Cette fonctionnalisation sert à caractériser un groupe d’atome qui chaque fois présent dans une structure moléculaire différente en modifie les propriétés chimiques de manière analogue.
Elle permet également de réprésenter les chaînes carbonnées en chimie organique. Le methyl a un atome de carbone quand l’éthyl en a deux.
Comparons maintenant la partie ester de la formule moléculaire de l’acétylcholine brute avec sa formule développée en 2D sur Wikipédia : C7,H16,NO2
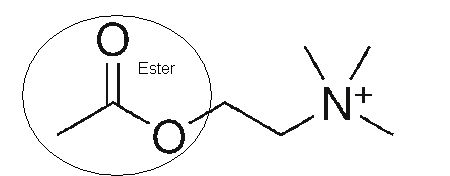
on retrouve la double liaison caractéristique entre l’atome d’oxygène et l’atome de carbone, l’ester d’acétate se caractérise par la chaîne O–CO–CH3 :
Ester D’acétate (Acétylisation de la choline ) –> AcetylcholineFormule semi développée de l’acétylcholine (Basé sur l’ion positif d’Azote de symbole atomique N)Représentation complète en 2 dimensions
Reprenons ou nous en étions :
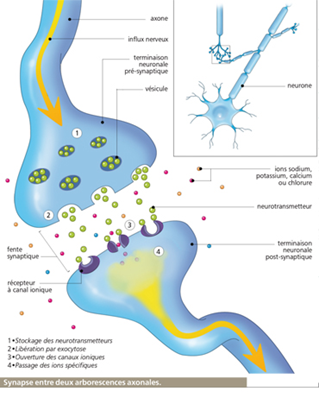
Schéma fléché de la libération de l’acétylcholine de sa synthèse par le nerf moteur jusqu’à sa destruction par une enzyme pompe, la cholinestérase.
Cette molécule est libérée par l’influx nerveux entre le nerf moteur et le muscle strié. A la différence de la synapse électrique, la synapse chimique est polarisée (directionnel) et l’espace entre le nerf moteur et le muscle strié est 10 à 25 (20 à 50 nm) fois plus grand que l’espace entre deux neurones électrique.
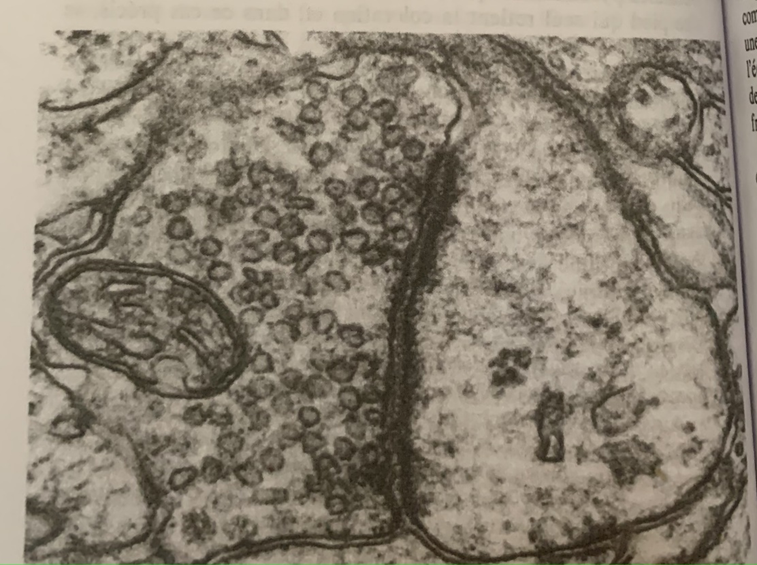
D’autre part, leur morphologie est différente, la synapse chimique est asymétrique : d’un côté une terminaison nerveuse bourrée de vésicules de 30 à 60 nm pleines du neurotransmetteur. De l’autre un épaississement de la membrane ou densité post-synaptique.
Grossissement de la synapse au microscope électronique 10^5. A gauche une mitochondrie ou métabolite énergétique de la cellule avec des multiples vésicules, à droite une épine dendritique dont la fonction morphologique est d’optimiser la liaison inter-neuronale. Au centre une synapse avec un épaississement post-synaptique caractéristique de son asymétrie.
Ces synapses révèlent la présence d’électricité générée par les impulsions issues de l’influx nerveux qu’elle transmet à la fibre musculaire de manière unidirectionnelle. Ces synapses fonctionnent comme une valve. Le signal ne fonctionne que dans un sens du nerf moteur au muscle.
On observe un délai entre 0,3 et 0,8 ms à la mesure de l’électricité dans le muscle, plus long que celui de l’influx nerveux : relation entre vitesse, distance et temps puisque le neurotransmetteur n’est pas présent dans le muscle à la base ce qui explique la polarisation.
Mécanisme de libération de l’acétylcholine
A la terminaison nerveuse, au niveau de la jonction nerf-muscle on sait que l’impulsion électrique provoquée par l’influx nerveux provoque la libération de 300 paquets d’environ 10.000 molécules chacune d’acétylcholine ce qui représente 3.000.000 de molécules.
Ce nombre est en définitive petit comparé à l’unité de matière basé sur la constante d’Avogadro qui est de 6,02 x 10^23 molécules dans une mole de matière mais dans un volume si petit que cette concentration est relativement forte.
Au repos, on a donc 10^-9 mole par litre d’acétylcholine mais lors du passage de l’impulsion nerveuse cette valeur est donc comprise entre 10^-4 et 10^-3 moles par litre pour une durée de l’ordre de la milliseconde puis s’évanouit grâce à une enzyme spécialisée : la cholinestérase.
Retour d’une onde électrique
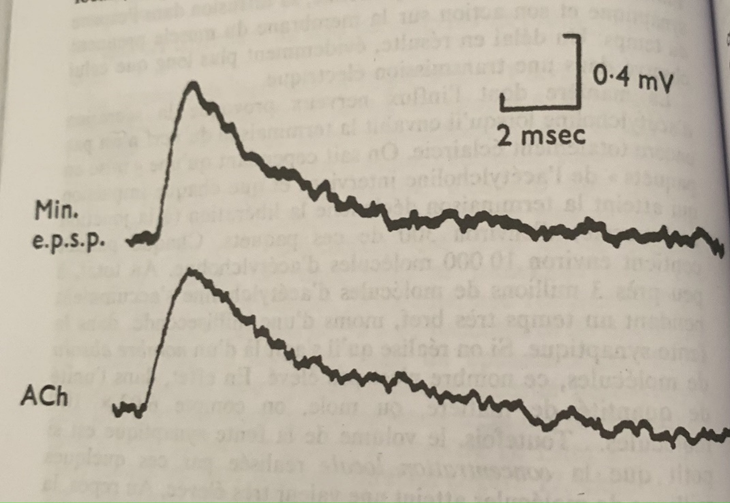
De l’autre côté de la terminaison nerveuse, au niveau de la face post-synaptique on enregistre une onde électrique bien différente de l’électricité générée à la base par l’influx nerveux. Son amplitude est 5 à 10 fois plus faible et sa durée est à 3 à 5 fois plus longue. (3 à 5 ms).
Réponse électrique de la membrane post-synaptique d’une synapse chimique (ici la jonction nerf-muscle) La forme et les propriétés de l’onde diffèrent de celles de l’influx nerveux. Le tracé du haut a été enregistré après la libération d’un seul paquet d’acétylcholine par la terminaison nerveuse. Celui du bas lors de l’application locale d’une goutte par micropipette d’acétylcholine. Les tracés se superposent.
Cette onde résulte de la perméabilisation de la membrane de la synapse qui engage les ions sodiums, potassiums et calciques individuellement et non séquentiellement. (Un canal s’ouvre individuellement sans lien commun avec les autres et brutalement (1ms) (le caractère individuel rend plus simple la transmission du signal)
En A l’activité électrique de la membrane sous synaptique de la face post-synaptique avec à gauche l’onde sans acétylcholine, à sa droite avec acétylcholine, Forte augmentation du bruit. (1 et 2) En B l’ouverture et la fermeture brutale unique (carré) des canaux sodiums, potassiums, calciums bien individualisés résultant de l’onde électrique générée par la transmission d’acétylcholine.
Résumé synthétique de la multiplicité des signaux
La libération de l’acétylcholine par l’influx nerveux entraîne la transformation d’un signal électrique (1) en signal chimique(2) L’ouverture des canaux ioniques constitue l’étape inverse et la transformation du signal chimique(3) en signal électrique(4) que j’appellerai onde de retour : elle résulte de l’ouverture de canaux au niveau de la synapse et est réencodée vers le premier neurone de la terminaison neurale pré-synaptique par lequel l’influx nerveux est passé afin de lui communiquer que le signal a été transmis.
Synthèse morphologique
Au niveau de la jonction nerf muscle strié, le potentiel déclenché par l’influx nerveux et la réponse synaptique dont résulte la transmission d’acétylcholine suffit à déclencher un influx. A chaque impulsion du nerf répond une contraction du muscle strié. La transmission est efficace à 100 %. Ce n’est pas toujours le cas en revanche pour des synapses entre deux neurones.
Ces terminaisons sont si petites qu’elles ne peuvent libérer qu’un seul paquet d’acétylcholine et pas nécessairement à chaque fois qu’un influx nerveux envahit la terminaison. Il n’en reste pas moins que le schéma général de la transmission chimique établi au départ avec la jonction nerf moteur muscle strié s’applique au système nerveux central.
Types de neurotransmetteurs
Franck Campbell McIntosh 1909-1992
L’acétylcholine a été isolé dans le cerveau pour la première fois par McIntosh en 1941 et la noradrénaline par Vogt en 1954 et ces deux neurotransmetteurs sont les membres d’honneurs d’une liste de substances chimiques qui s’allongent de jour en jour tels que :
Acides aminés
Amines biogène « double fonction » : dopamines, sérotonine…
Ce dernier a été découvert dans l’intestin ce qui montre bien que l’interaction entre le système nerveux périphérique et le système nerveux central.
Un autre exemple est la somatostatine qui bloque l’hormone de croissance et qu’on retrouve dans le pancréas.
On retrouve ces neurotransmetteurs en particulier chez tous les vertébrés. Aucun neurotransmetteur n’est propre à l’être humain. (Chimie prégnante dans le vivant)
La diversité biochimique qui ne cesse de s’accroître au fur et à mesure des recherches en neurosciences contraste avec l’uniformité du signal électrique. Un même neurone peut recevoir des dizaines de milliers de synapses et que ces synapses peuvent employer des neurotransmetteurs différents.
Cette diversité dans la nature chimique et dans les effets ioniques introduit une combinatoire de signaux qu’un neurone électrique n’effectuerait pas de la même manière ou n’effectuerait même pas du tout.
Les molécules serrures
La synapse chimique tient une place centrale dans la communication entre deux neurones, Elle canalise le transfert de signaux de cellule à cellule en créant une polarité dans ce transfert, crée des circuits et introduit une diversité au niveau de la membrane du neurone. Ses dimensions sont de l’ordre de grandeur d’une cellule bactérienne.
Afin de démonter le fonctionnement de la synapse chimique, il convient d’étudier l’effet du neurotransmetteur sur la face post-synaptique. Hors Emil Fischer postule en 1906 que pour qu’une molécule puisse agir sur un corps, elle doit pouvoir se fixer.
« Corpora Non Agunt nisi fixata »
Emil Fischer 1852-1919
Comment cette fixation se fait-elle ?
La cellule contient une substance chimiquement active (un récepteur) qui porte une configuration géométrique adapté complémentaire de celle du corps considéré (le neurotransmetteur). La métaphore est celle de la clé dans une serrure.
Langley montre que cette substance répond au curare (effet inhibiteur) à l’acétylcholine et à la nicotine de manière analogue et qu’elle n’est localisée sur le muscle adulte qu’au niveau de la terminaison nerveuse : c’est la serrure ou la molécule réceptrice.
John Newport Langley 1852-1925
Schéma illustrant tout le fonctionnement d’une communication synaptique avec passage et nouveau déclenchement d’influx nerveux (macromolécule : réceptrice en violet qui a la forme de micro sacs adaptés à la réception des différents ions dont l’ouverture de canaux pour le retour d’une onde électrique séquentielle. (Vu plus haut)
Comment le prouver ?
La nature recèle de richesse et le poisson « torpille » aussi appelé « gymnote » dont trois décharges de 500 V à 0,5 A tuent un homme résultent de la mise à feu d’un milliard de synapses très semblables par leurs propriétés à celles que l’on trouve à la jonction nerf-muscle de l’humain. L’organe électrique se compare à un muscle dont l’appareil contractile aurait été enlevé pour laisser les synapses à vif.
Cette accumulation gigantesque va se révéler fort utile pour le biochimiste. En plus ces synapses ont toute la même composition chimique : travailler sur 1 kg d’organe chimique revient à travailler sur une synapse géante qui pèserait le même poids.
Poisson Torpille
Dans un premier temps on va utiliser comme produit radioactif pour marquer la face post-synaptique et détecter la forme et la position de cette fameuse molécule réceptrice le neurotransmetteur lui-même (acétylcholine). L’expérience échoue car de nombreuses molécules lie de manière non sélective à beaucoup trop de molécules du récepteur lui-même.
La nature va encore venir au secours des biochimistes : les serpents cobra et bungare ont une mauvaise réputation de par la toxicité de leur venin qui paralyse les voies respiratoires au même titre que le curare des Amérindiens. Cette toxine alpha qui bloque les récepteurs va être utilisé comme marqueur radioactif pour détecter la localisation sur de la molécule en question.
Serpent Bungare
On trouve des serpents marins mangeurs de poissons torpilles dans la mer du Japon, leur réunion dans le tube à essai va permette d’isoler cette fameuse molécule réceptrice sur laquelle se fixe l’acétylcholine.
Cette molécule isolée a une masse de 250.000 soit 3,5 fois plus que celle de l’hémoglobine, elle se compose de plusieurs chaînes de quatre types différents dont une répétée deux fois. Elle est disposé sur toute la membrane sous-synaptique de la face post-synaptique.
Leur forme géométrique étudiée par centrifugation de broyats d’organes électriques est celle de micro sacs que l’on peut remplir d’ions sodiums ou potassiums radioactifs pour mieux comprendre l’effet de l’acétylcholine qui provoque l’ouverture des canaux.
Ou se trouvent ces canaux ioniques ? On disperse les micro-sacs grâce à des détergents sans les détruire dans le tube à essai. Elle garde ainsi toutes ses fonctions natives comme sur la membrane sous synaptique. On peut ainsi en étudier son électrophysiologie qui renvoie plusieurs signaux exactement similaires à l’ouverture de plusieurs canaux individualisés brutalement (1 ms) de forme carré.
La molécule de masse 250.000 contient à la fois le site récepteur et le canal ionique.
Schéma traduisant l’expérience de la synapse géante du poisson torpille qui prouve l’ouverture brutale du canal ionique et le passage du sodium Na++ localisé sur la molécule réceptrice. (Flèche)
Cette molécule est analogue à une protéine allostérique, ce concept défini par Jacques Monod et Jean-Pierre Changeux veut que la fixation d’une molécule effectrice sur un site modifie les conditions de fixations d’une autre molécule en un autre site distant de la même protéine un peu comme deux particules quantiques intriquées.
Ces protéines sont sujettes à des transitions discrètes entre états moléculaires distincts actifs ou inactifs. Dans notre cas, le récepteur, l’illustration est l’ouverture et la fermeture ultra-rapide des canaux. #carré
L’onde post-synaptique enregistrée au niveau de la fibre musculaire correspond donc à l’ouverture simultanée d’une collection importante de récepteurs-canaux. #fibre
Elle s’identifie à la somme d’évènements moléculaires discrets au même titre que l’impulsion nerveuse s’identifie à l’ouverture de canaux sodiums dans la membrane de l’axone et aux flux ioniques qui en résultent.
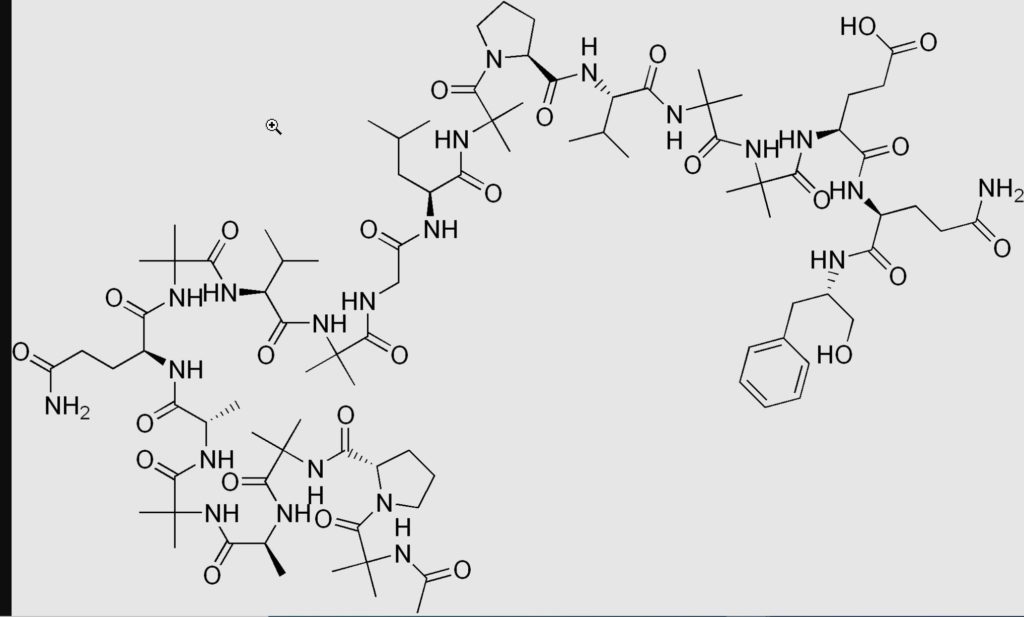
La molécule réceptrice comme toute protéine cellulaire se compose d’acides aminés enchaînés les uns aux autres (environ 2500) Chaque acide aminé contient entre 10 et 30 atomes qui déterminent intégralement sa réactivité chimique, ses possibilités de liaison avec ses congénères (polymérisation) (plusieurs parties réunies) pour construire son édifice protéique (macromolécule) (Ex : peptide)
@Wikipédia Comparez la structure moléculaire d’un seul acide aminé avec celle d’un peptide et vous comprendrez la polymérisation et la constitution d’une protéine que l’on peut qualifier de manière réductrice de macromolécule.
Conclusion transitionnelle
Nous accédons donc à une organisation au niveau moléculaire, atomiques et ioniques de la transmission chimique entre deux neurones.
Synthèse
Les atomes psychiques réexaminées (Démocrite)
La même méthode en définitive, conduit de l’enregistrement de l’encéphalogramme à celui des impulsions électriques produites par des neurones uniques, de la réponse post-synaptique aux transitions d’ouvertures des canaux ioniques, eux même identifiables à des changements de structure moléculaire.
A chaque étape il y’a réduction d’un niveau d’organisation à une unité plus élémentaire : de la population des neurones corticaux, à la cellule ou à la synapse, de l’impulsion nerveuse (ou à des courants post-synaptiques) à la molécule.
L’étude dynamique des différentes pièces du cerveau sont donc descriptibles avec les mêmes termes que ceux du physicien et du chimiste.
On pourrait plus loin et décrire la formule atomique de la molécule réceptrice dont on vient d’étudier le fonctionnement pour comprendre sur quoi se base sa réaction à l’acétylcholine.
L’activité spontanée et évoquée est donc directement issue de propriétés atomiques. Doit-on alors réexaminer les atomes psychiques de Démocrite ? Les ions sodiums et potassiums qui traversent les canaux de l’axone sont les mêmes que ceux que l’on trouve dans l’eau de mer ou à l’intérieur du neurone.
Les molécules de neurotransmetteurs sont composées de carbone, d’hydrogène, d’oxygène et d’azote qui n’ont rien de propre aux êtres vivants. Le système nerveux se compose de et emploie pour fonctionner – la même matière que le microcosme.
Celles-ci s’organise en édifices moléculaires qui interviennent dans la communication nerveuse au même titre que d’autres règlent la respiration cellulaire ou la réplication des chromosomes. Les protéines tiennent la une place critique puisqu’enzyme pompes (destruction ou aspiration), enzymes de synthèse des neurotransmetteurs et leurs récepteurs sont des protéines.
Au lieu de reprendre le terme d’atomes psychiques, doit- on parler de molécules psychiques ? Le trait le plus frappant qui se dégage des recherches actuelles, c’est que l’électricité et la chimie du cerveau sont les mêmes que celles des organes périphériques. On retrouve dans la nature : ce qui est vrai pour l’organe électrique du Gymnote est aussi vrai pour Homo Sapiens.
Au niveau des mécanismes élémentaires de la communication nerveuse, rien ne sépare l’homme des animaux. Aucun récepteur, canal ionique ou neurotransmetteurs n’est propre à l’homme.
Cet article est un résumé du chapitre 2 du livre « l’homme neuronal » de Jean-Pierre Changeux neurobiologiste célèbre qui a révolutionné les pistes de recherches dans le domaine des neurosciences.
Jean Pierre Changeux 1936 – X
Partons du général pour arriver au plus précis :
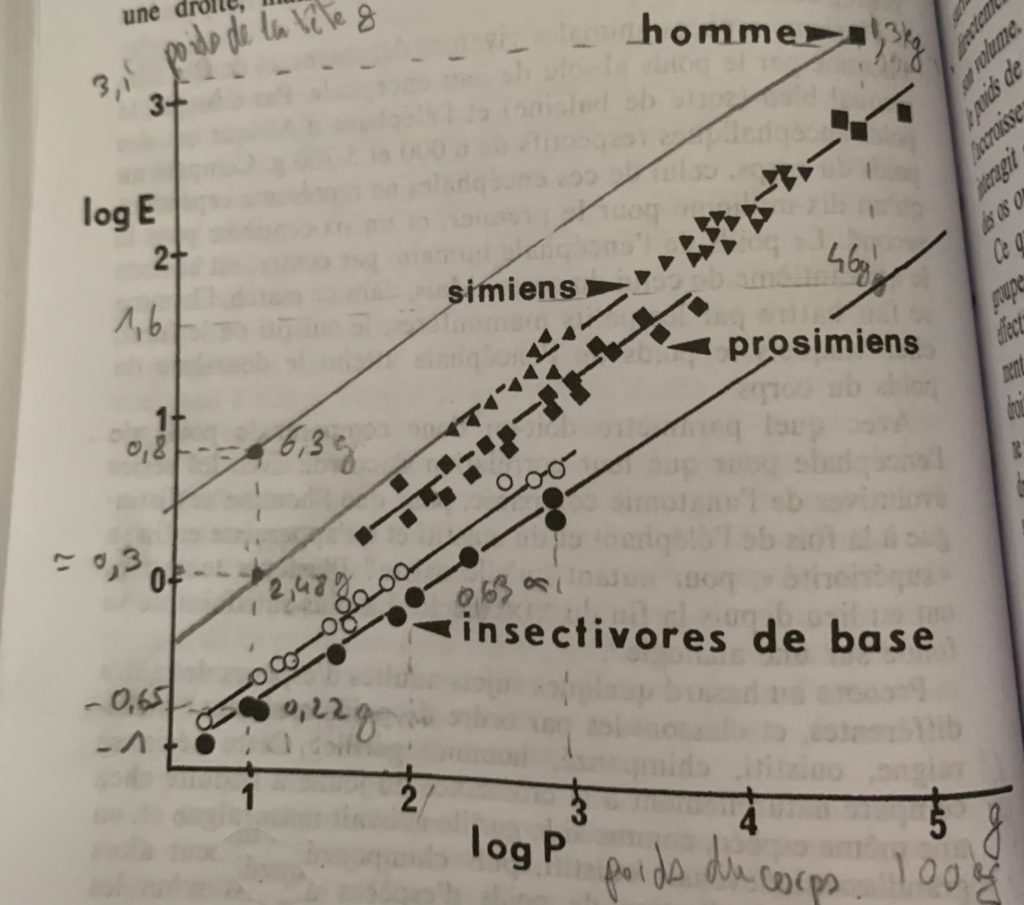
Un classement a été établi d’après Roland Bauchot et Heinz Stephan afin de faire un parallèle entre le poids du corps et le poids de l’encéphale de plusieurs espèces par le calcul d’un indice d’encephalisation.
Lorsque l’on applique des coordonnées logarithmiques (dont le résultat est basé sur le nombre de puissance) avec une fonction exponentielle (fonction puissance elle-même) on obtient une fonction linéaire ce qui est très pratique pour comparer les évolutions entre différentes espèces. On obtient ainsi une série de droite parallèles qui ne se superposent pas.
Echelle ou système de coordonnées logarithmique avec le poids du corps en abscisse et le poids de la tête en ordonné. La fonction exponentielle est la bijection réciproque de la fonction logarithme décimal ce qui explique pourquoi on obtient une fonction linéaire. @Bauchot et Stephan
Explication arithmétique et pédagogique de la représentation graphique :
Pour que vous puissiez remonter sur le graphique au fur et à mesure des explications je vais vous créer des ancres hypertextes internes tout du long de celles-ci.
J’ai glosé et calculer tous les poids, tracer des lignes pour les correspondances en fonction des logarithmes, mais à titre pédagogique exerçons nous.
Si vous avez suivi mon article sur les logarithmes la base employée ici est 10, quelle valeur le résultat d’un logarithme égal à 5 pour un nombre de puissances de 10 ? :
100.000 grammes soit 10^5 soit 10^2 kg de poids de corps. #graphique
Vous remarquerez que mon crayon à papier a volontairement tracé la ligne de l’homme à partir du point (elle n’est pas tracée sur le schéma original, ainsi que la continuité des singes (simiens) car l’auteur fait une comparaison unitaire entre les trois espèces :
Pour un logarithme du poids du corps égal à 1 entre les trois espèces soit 10g le singe a un cerveau environ de 11,3 plus gros que celui de l’insectivore de base. L’homme a un cerveau 28,7 fois plus gros. #graphique
Fonction antilogarithme
Il parle aussi d’un insectivore avec un poids égal à logarithme de 5 alors j’ai fait un point pour représenter un insectivore avec un tel poids et faire une correspondance en pointillé avec le poids de l’encéphale en ordonnée. #graphique
Partez toujours de l’abscisse soit log = 5, prenez votre doigt et remontez jusqu’au point de l’homme, par lecture graphique on voit que la correspondance avec l’ordonnée est log = 3,1. #graphique
Comment faire pour calculer le poids exact en kg ? je vais introduire un autre concept du logarithme fort simple, la fonction antilogarithme. On sait que la base utilisée est 10 et on sait que le résultat du logarithme est égal à 3,1 approximativement d’après notre tracé horizontal en pointillé. #graphique
On va utiliser la réciproque du logarithme décimal ou antilog que j’ai formalisé sur LATEX (voir mon article dessus) #graphique
Pour l’anecdote j’ai trouvé un code source latex qui symbolise l’antilogarithme comme pouvez le voir, il a la forme d’un N inversé qui s’étend comme une racine
La pente
La pente d’une droite est son inclinaison en ordonné pour un espace unitaire entre deux valeurs de x, par lecture graphique vous pouvez voir qu’elle est égale à 0,63. #graphique
Sur le schéma elle est représentée par les deux tracés pointillés verticaux entre 2 et 3 de log Poids du corps. Cette valeur alpha α correspond au rapport entre la surface et le volume dans cette représentation ce qui veut dire que le poids de l’encéphale est plutôt indexé sur la surface du corps que sur son poids.
Conclusion
L’homme dépasse donc tous les vertébrés, le chimpanzé est proche (moins d’un facteur 2,5 soit : 520 g
Log (520) = 2,7 plus dangereux sont les phoques et les dauphins dont le coefficient par rapport à l’insectivore de base est respectivement de 15 et 20. #graphique
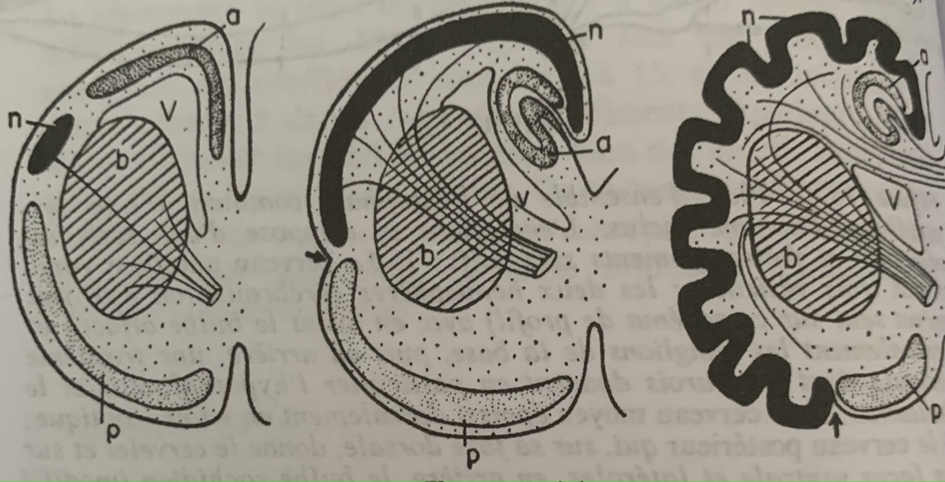
Annexe partie 1 : Expansion du néocortex entre 3 vertébrés
Bauchot et Stephan compare maintenant l’évolution du cortex à l’intérieur de l’encéphale entre les reptiles, les primates et l’homme grâce à un indice de progression, le reptile est à 1, le primate qui a le néocortex le plus développé est à 58 (chimpanzé) tandis qu’homo sapiens à un indice de 156.
Heinz Stephan 1924-2016
Le néocortex est si développé qu’il occuperait dans le cas où le cerveau aurait la forme d’un cube 7 dm² alors que la surface totale de l’écorce cérébrale mesure 22 dm². On connaît donc le pourquoi de la morphologie des circonvolutions du cerveau en plicatures enfouies dans les sillons en profondeurs.
En noir le néocortex (à gauche insectivore, au centre le chimpanzé, à droite l’homme.)
Chez l’homme, la fraction de l’encéphale représente une part de plus en plus importante du poids du corps. Cela s’explique en partie par la corticalisation de l’encéphale qui est une caractéristique de tous les mammifères. (A noter qu’entre les trois schémas, le bulbe olfactif représenté par a ne fait que diminuer) L’homme à moins d’odorat que beaucoup d’animaux primitifs.
Partie 2 : Typologie et topologie des cellules du cortex
Dans le Cortex Baillarger grâce à des coupes histologiques a pu étudier le type de cellules présentes dans un cortex qu’il stratifie en 6 couches. Chez l’homme on compte deux catégories principales de cellules inégalement réparties à travers son épaisseur :
François Baillarger 1809-1890
François Baillarger
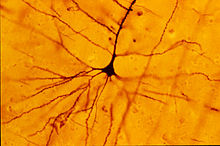
Les neurones se différencient donc en neurones pyramidaux qui se caractérisent par la longueur de leur axone et la forme de leur corps cellulaire comme leur nom l’indique. Les cellules en étoiles ou astrocytes font partie des cellules glies (vient du grec « gloios » qui veut dire glue)
Quant à ce qu’on appelle les neurites: pour faire simple ce sont les axoneset les dendrites.
Les dendrites pour un neurone peuvent être apicales (au sommet de l’axone) ou basales (à la base du corps cellulaire).
Les cellules étoilées ont des corps cellulaires(soma) plus petits et de structure ovoïde. A titre d’exemple, dans l’aire visuel du cortex elles se répartissent dans trois couches densément et ont même exceptionnellement des épines au bout de leurs dendrites.
Les cellules étoilées ont des axones qui se projettent moins vers la surface et ont une fonction d’interneurone avec les cellules pyramidales caractérisée par leur arborisation dendritique différente ( en bouquet) de celle des cellules pyramidales.
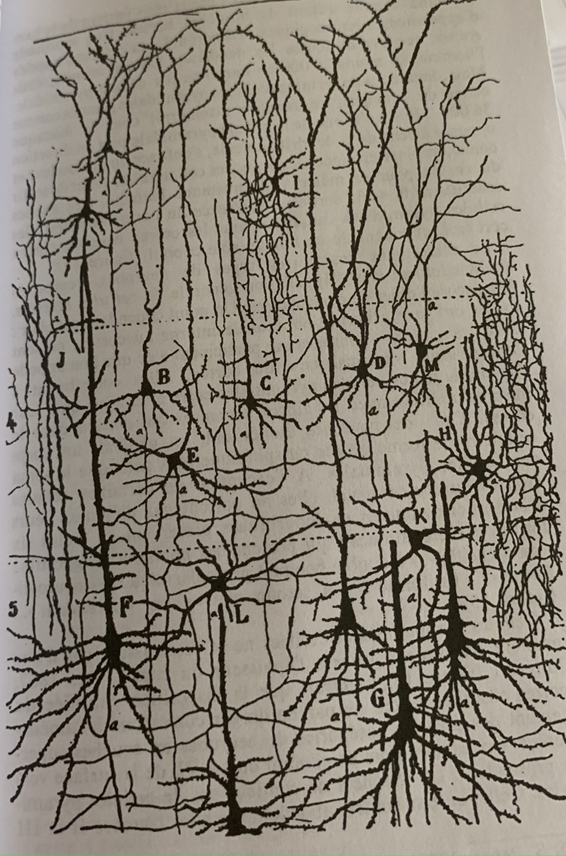
Les cellules F, A, G, B sont des cellules pyramidales, elles projettent leur axone vers la surface du cortex tandis que les cellules étoilées C, D, E, H, M restent à l’intérieur et assurent le rôle de soutien. Elles ont des dendrites ramifiées de différentes sortes : à double bouquet (H), à axone court et ramifié en longues branches horizontale (E) etc…
Même si à travers les cortex ces deux types de cellules se répartissent différemment globalement l’homme et les mammifères ont un cortex avec une grande unité morphologique.
En effet à travers le lobe pariétal, temporal, occipital, frontal, on trouve la même catégorie de cellules qui seront spécifiées plus tard par les interactions biochimiques (neurotransmetteurs).
Le nombre de cellules pour une carotte entre l’homme, le primate et le rat est le même sauf dans l’aire visuelle de tous les mammifères ou l’on trouve 2,5 fois plus de cellules ce qui est logique compte tenu du volume et de la richesse d’informations transmises à traiter par notre rétine.
On en revient donc à la surface du cortex et de manière indu à l’expansion du néocortex comme facteur d’accroissement du nombre de neurones et de calculs chez l’homme.
L’architecture cellulaire du cortex varie d’apparence d’une aire à l’autre. A gauche section à travers un cortex sensoriel (aire visuelle) où les cellules étoilées s’accumulent dans trois couches denses d’où le nom de cortex granulaire. A droite cortex moteur avec des cellules pyramidales de grande taille.
Par contre chez les invertébrés on compte une grande diversité de ces catégories de cellules ce qui rend le fonctionnement de leur cortex plus complexe à étudier, à l’exception des céphalopodes qui ont la même tendance évolutive que les mammifères.
Partie 3 : Câblage
Grâce à la révolution de la microscopie électronique, on aurait pu croire qu’il aurait été plus facile d’étudier comment ces neurones sont reliés mais l’uniformité de la synapse est encore plus grande que celle des cellules.
Comment faire alors sachant qu’il y’a plus de 10^15 synapses dans le cortex de l’homme ? Cela prendrait 30.000 ans de les dénombrer une par une.
Powell a réussi à voir qu’il y’avait le même pourcentage de cellules en étoile et de cellules pyramidales dans deux aires très distinctes du cerveau le cortex moteur et le cortex visuel. Même si leur distribution dans les différentes strates comme on l’a vu est inégale. On peut donc schématiser comment le câblage s’opère.
Thomas Shroud Powell 1923-1996
Entrée/Sorties
Pavlov a prouvé que les organes des sens se projettent au niveau de certaines aires spécialisées, pourtant aucun d’entre eux n’entrent directement dans le cortex. Ils empruntent plutôt la voie du thalamus dans lequel certains neurones vont prendre le relais au niveau d’aires sous-corticales.
Ivan Pavlov 1849-1936
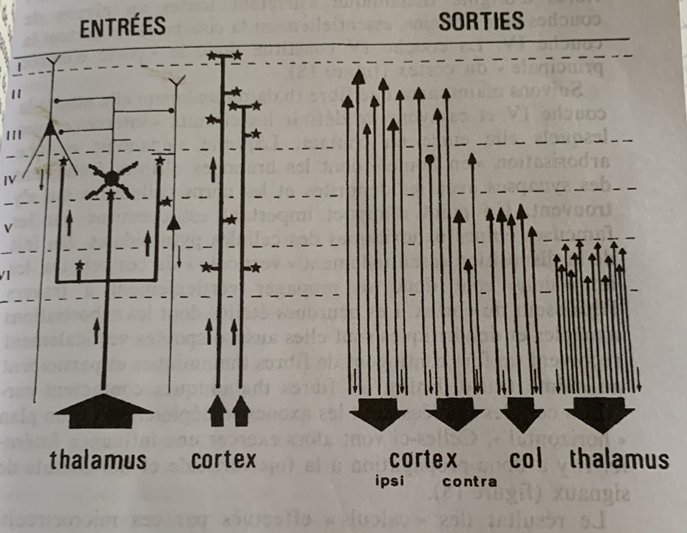
Représentation des entrées et des sorties ainsi que des interconnexions entre les différentes types de cellules. les lignes : les fibres de neurones avec leurs axones, Les étoiles : représentent les synapses, les chiffres romains : les couches corticales, les traits horizontaux les connexions avec les cellules gliales ou astrocytes.
Pour mieux comprendre le schéma et son explication je vais créer des ancres ou hyperliens à l’intérieur de la page pour vous permettre de remonter et de regarder le schéma en parallèle du texte :
Toutes les aires corticales (6 couches en chiffres romains) qu’elles soient motrices ou d’association reçoivent des fibres provenant du noyau thalamique dans une unité d’organisation remarquable. Le cortex lui-même en est un point d’entrée. (2ème en partant de la gauche).
Les fibres issues du thalamus s’arrêtent toutes à la couche IV dans une arborisation en boule ou les synapses (étoiles) vont se relier aux dendrites ou d’autres neurones vont prendre le relais. Les impulsions vont donc se propager verticalement mais aussi par contact horizontal avec les astrocytes (lignes horizontales pointées). #schéma
On a pu analyser grâce à la coloration au peroxydase du raifort que la sortie des entrées du cortex est le cortex lui-même (3ème en partant de la gauche) dont certaines fibres d’association retournent vers le cortex du même côté (couches en chiffres romains) (ipsi) ou du côté opposé (contra) pour former des boucles. Le reste des sorties des cellules pyramidales s’arrêtent dans des zones sous-corticales. #schéma
Le second point d’impact est le noyau du thalamus (5ème en partant de la gauche) dont là encore les fibres du cortex retournent dans le cortex pour former un autre circuit en boucle, (les calculs s’enchaînent) #schéma
Enfin le troisième et dernier point d’impact des cellules pyramidales sont les autres aires sensorielles telles que le Colliculus supérieur qui coordonne les mouvements des yeux et de la tête. Celles-ci vont déclencher des réactions motrices. Mais certains axones vont encore plus loin jusqu’à la moelle épinière dans des neurones moteurs qui commandent les contractions musculaires. #schéma
Ainsi on peut effectuer une analogie entre le fonctionnement du cerveau et celui de la machine dont les entrées-sorties peuvent former des boucles. #schéma
Partie 4 : Modules ou cristaux
Modules
La structure des interactions entre chaque neurone est-elle organisée en module ce qui implique une fixité des dimensions (géométriquement définies) et répétés ? Vernon Mountcastle va reprendre les techniques d’encéphalogramme de Caton (voir mon article sur l’histoire des neurosciences) qu’il va optimiser à l’aide d’électrodes de plus petite taille qu’il va implanter dans la zone corticale pour étudier les réactions des neurones chez le chat sur un plan transversal.
Vernon Mountcastle 1918-2015
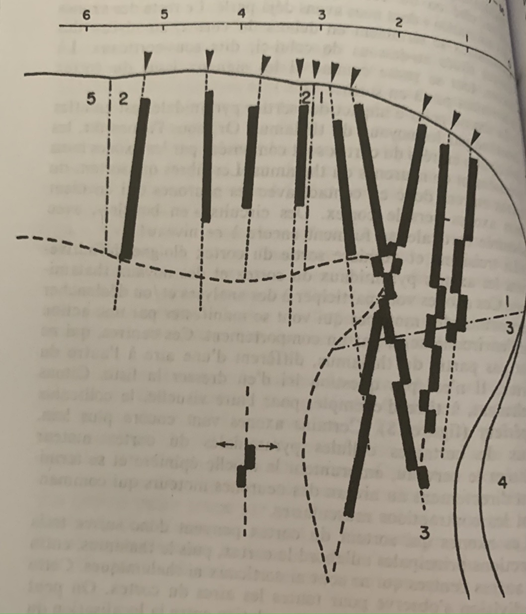
Lorsqu’il implante l’électrode dans l’alignement vertical les bandes de neurones répondent selon la même modalité (verticalement). Lorsque l’électrode est implantée sur un plan oblique la modalité varie brusquement ce qui laisse supposer que les colonnes sont organisées non par leur géométrie verticale dans le cortex mais par leur organisation avec les périphéries qui constitueraient les modules du cortex.
Les petites flèches sont les électrodes au centre placé en ligne droite, les trois sur la droite sont oblique ce qui montre une variation de la modalité représentée par la discontinuité des bandes verticales en noir.
Hubel et Wiesel vont aller plus loin, même si dans un premier temps la stimulation sensorielle à partir de chaque œil semble confirmer l’organisation verticale et la fixité de dimensions propre à une structure modulaire, lorsqu’ils vont changer de plan et voir le cortex de haut c’est-à-dire à plat, ils vont remarquer un réseau similaire aux bandes noires et blanches d’un zèbre.
Torsten Wiesel 1924-X
Dave Hubel 1923-2013
En enlevant complètement l’œil d’un chat à la naissance ils vont même voir que les bandes de l’œil qu’ils ont gardé envahissent toute la structure corticale par compensation, les bandes œil droit, œil gauche n’ont donc pas de dimensions fixes.
Les points de tissage corticaux du thalamus forment une arborisation en boule comme nous l’avons dit précédemment et s’organisent donc selon un tissage en 3 dimensions. En fait l’organisation neuronale du cortex est donc extrinsèque (part du thalamus) et non intrinsèque.
De même lorsque l’on stimule l’œil du chat avec de la lumière les bandes n’ont pas la même taille que par la stimulation de l’électrode. Le schéma en module de dimensions fixes même avec des petits modules dans des hyper modules emboîtées est donc trop réducteur.
Cristaux
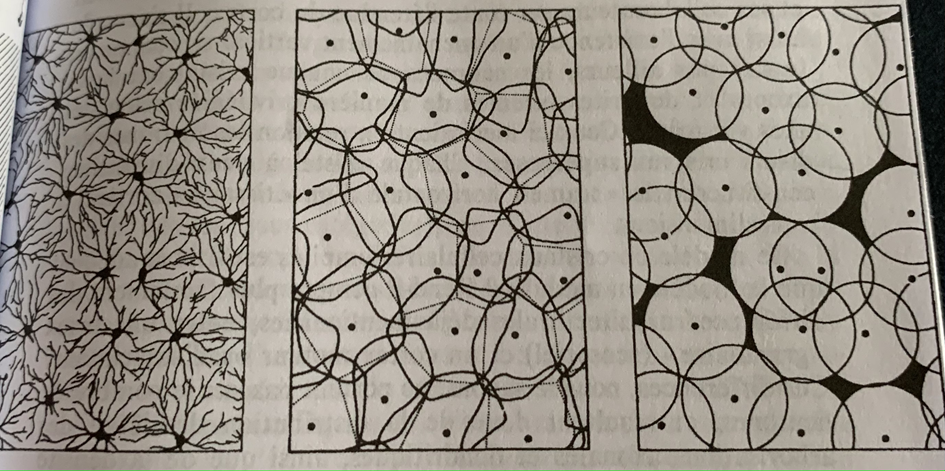
On va chercher d’autres aires cérébrales dont la disposition cellulaire est plus simple et notamment le cortex du cervelet (visuel et granulaire) que l’on connaît bien. Chez les vertébrés la typologie de cellules est la suivante :
On compte 5 types de cellules dont celles de Purkinje qui peuvent être prises comme homologues des cellules pyramidales, les cellules en grain comme l’analogue des petites cellules étoilées et le reste est composé de 3 gros types de cellules étoilées.
Dans le cervelet les cellules s’organisent en trame horizontale dans le plan du cortex, elles s’organisent comme un nœud de réseaux, les dimensions varient d’une catégorie de neurones à l’autre. Une organisation analogue se trouve dans la rétine. Les hypothétiques modules s’organisent en fait sous la forme de cristaux en deux dimensions dont l’interconnexion en forme une troisième.
Distribution régulière des cellules ganglionnaires dans le plan de la rétine : à gauche cellules étoilées, au centre schématisation ou leurs connexions sont tracés par un trait continu. A droite, schématisation en cercle ou la régularité paraît moins bonne qu’au centre. Les cellules d’une même catégorie s’organisent de manière quasi cristalline.
Cette organisation en cristal est adaptée aux système d’entrée/sorties que nous avons vu et l’arborisation en boule du thalamus (couche IV du schéma entrée sortie), d’autre part, plus l’axone est long plus le corps cellulaire est gros et par la même occasion les dendrites basales ce qui contredit l’hypothèse en module de dimensions fixes.
Par ailleurs, on sait que chaque aire corticale est différente en termes de disposition et de densité cellulaire ce qui singularise chaque connexion cristalline.
Notre cerveau est donc un répertoire de singularités de connexions reliées par des dizaines de milliers de synapses. La tâche pour comprendre chaque singularité est immense !
Synthèse
Le cerveau de l’homme ne diffère en rien de celui des mammifères dans leurs typologies et leurs morphologies. En effet dans une carotte de cortex on trouve la même densité de cellules et de synapses du rat à l’homme.
Cependant le cerveau de l’homme se distingue donc par l’expansion du néocortex, son épaisseur trois fois plus grande, mais aussi principalement par l’augmentation de sa surface 400 fois par rapport au rat. Celle-ci reflète un saut quantitatif résultant de l’accroissement de nombre de connexions possibles.
D’autre part, chaque aire du cerveau humain est différente de par la disposition et distribution intrinsèque aux zones corticales des deux types de cellules dans les différentes strates ce qui augmente de manière exponentielle les fonctions de calcul et sa singularité.
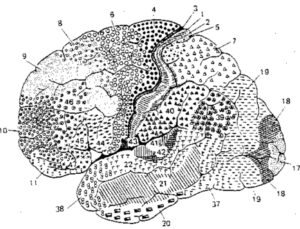
Afin d’introduire notre catégorie sur les neurosciences, il me paraît opportun de vous faire part de leur histoire au fil des siècles de l’antiquité jusqu’à aujourd’hui. J’ai utilisé beaucoup d’images de portraits volontairement pour travailler votre physionomie ainsi que des illustrations d’appareils cérébraux pour rendre le propos plus concret.
Les sciences du cerveau ont impliqué à travers l’histoire un nombre conséquent de grands hommes et femmes qui ont cherché à comprendre une première question : qu’est ce qui génère notre pensée ?
Premier débat sujet à controverses :Cœur ou cerveau ? Cardio-centristes contre Céphalo-centristes.
Déjà à l’époque comme dirait Jean Didier Vincent : on sait qu’il y a deux manières radicales de tuer un homme : lui couper la tête, ou lui planter une épée dans le cœur.
Démocrite (-460 -370) père de l’atome pense déjà que le siège de notre cerveau est dans notre tête. Pour lui notre cerveau serait constitué d’atomes psychiques qui parcourent tout notre corps. Ces atomes permettent d’animer nos muscles.
Démocrite
Aristote (-384 -322) s’oppose à cette idée, le cœur est pour lui le siège de nos fonctions motrices et psychiques. Pourquoi ? En anatomie, lorsque l’on envoie un fluide avec une pression sur le cœur, cela entraîne une réaction de contraction mécanique contrairement au cerveau.
Aristote
Le premier débat et qui durera fort longtemps était né et voit s’opposer céphalo-centristes aux cardio-centristes.
Galien (129-201) céphalo-centriste comme Platon, Hippocrate, Hérophile, et Erasistrate qui sont trois médecins grecs, va à peine plus loin que Démocrite. Pour lui l’âme est matérialisée par un pneuma (souffle) psychique qui circule du cerveau jusqu’à nos nerfs.
Galien 129-201
Erasistrate -310 -250
Hérophile -320 – 250
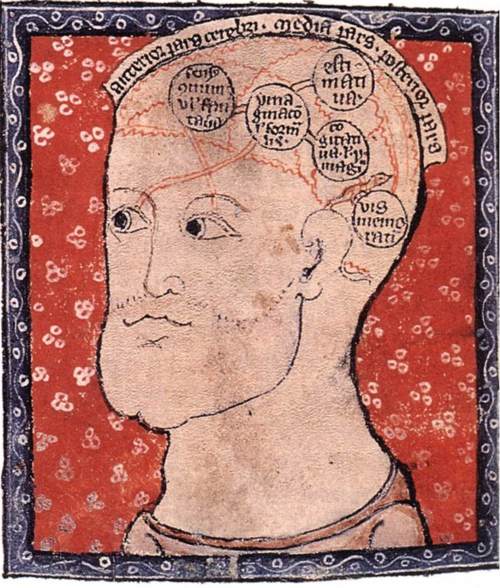
Les pères de l’église et notamment Némésius (350-420) vont plus loin, ils vont assigner des fonctions de notre pensée à des ventricules de notre cerveau. Ce sont les premières démarches localisationnistes.
Némésius
Cartographie des premières localisations de fonctions cérébrales par Némésius
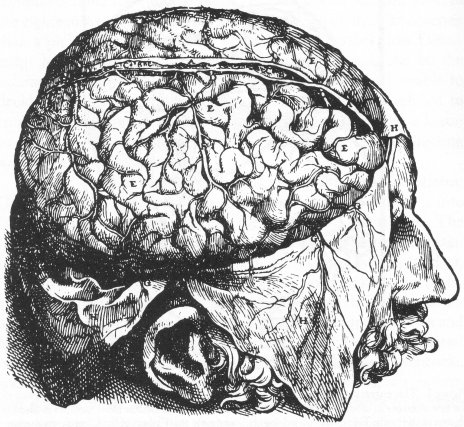
Au 16ème siècle au cours de la Renaissance va s’effectuer le retour des thèses anatomiques. Léonard de Vinci et Varole dissèquent le cerveau tandis que Vésale va illustrer avec fidélité toutes les circonvolutions et vaisseaux sanguins de notre appareil cérébral. Ils sont les premiers phrénologues.
Léonard De Vinci 1452-1519
Vésale 1514-1564
Varole 1543-1575
Dessin des circonvolutions exactes du cerveau par Vésale
Descartes (1596-1650) quant à lui va se montrer plus radical, pour lui le corps est une machine composée d’os de nerfs et de muscles, esprit plus rationnel qu’anatomiste, il va lier l’âme à la glande pinéale qui est unique mais réalisera une description du fonctionnement des mouvements du corps si précise qu’elle est encore en adéquation avec les théories actuelles sur l’arc réflexe.
René Descartes
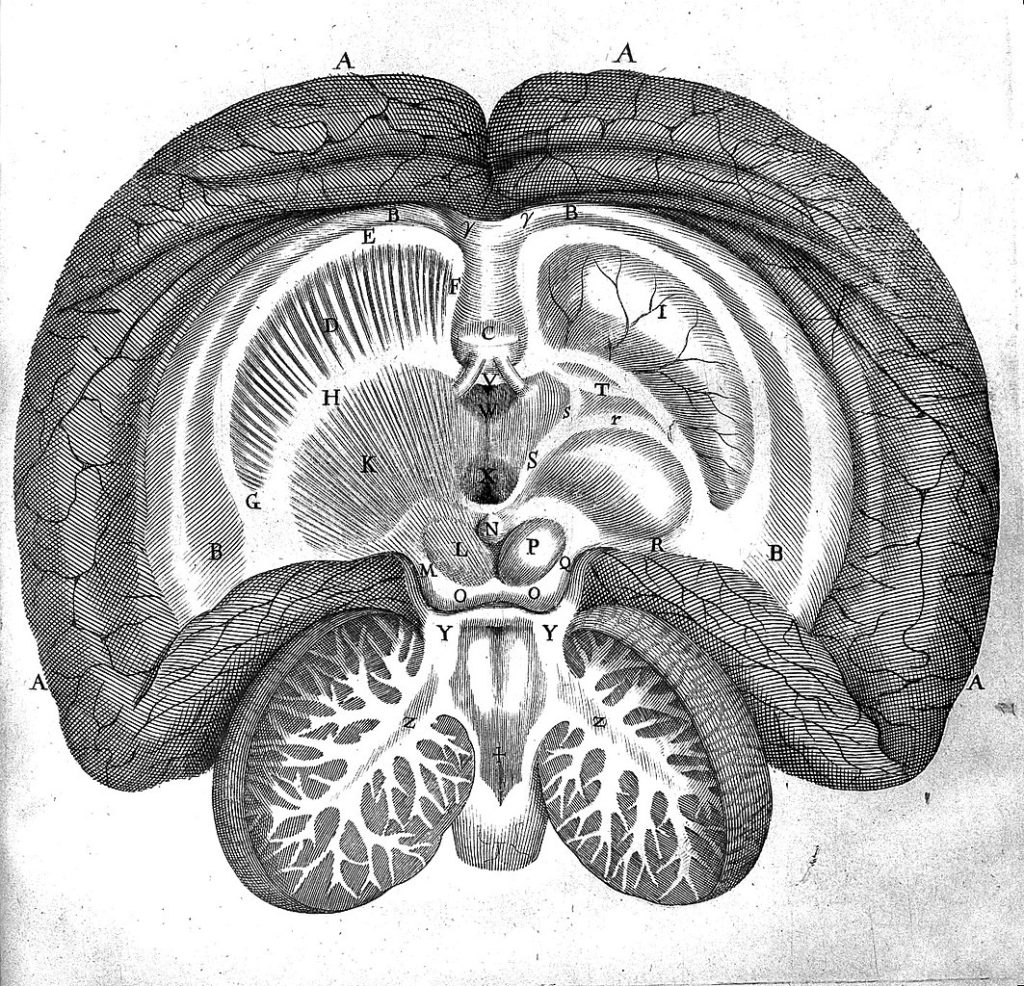
Thomas Willis (1621-1675) attribue la primauté au cortex cérébral, il identifie le corps calleux qui sépare les deux hémisphère et isole la matière grise et la matière blanche médullaire (moyen) dont la description est proche de celle qu’on utilise aujourd’hui pour la production d’influx nerveux. Cependant encore une fois pour ne pas s’attirer les foudres de l’église, il va lier l’âme aux corps striés mais accepte l’idée de son caractère immatériel.
Thomas Willis
Découpe du corps calleux en deux hémisphères et isolation de la matière blanche et matière grise par Willis
La fin de l’immatérialité de l’âme
En parallèle Gassendi (1592-1650) qui pourtant fait partie de l’église réhabilite les atomistes grecs et considère que les animaux qui font preuve de mémoire doivent eux-aussi avoir une âme ce qui dévalue beaucoup la supériorité de l’homme sur les animaux.
Gassendi
Le concept d’immatérialité de l’âme va être enterré peu à peu lorsque Vaucanson (1709-1782) dont j’ai déjà parlé dans mon article sur les sciences cognitives aidé par le chirurgien Le Cat (1700-1768) va fabriquer les premiers automates d’animaux avec le canard qui bat des ailes, se déplace, et digère les aliments. La Mettrie (1709-1751) se verra banni par l ‘église pour avoir dit que les hommes sont des animaux machines. Et Cabanis (1757-1808) dira que le cerveau secrète la pensée comme le foie secrète la bile.
Vaucanson
Le Cat
La Mettrie
Cabanis
La phrénologie
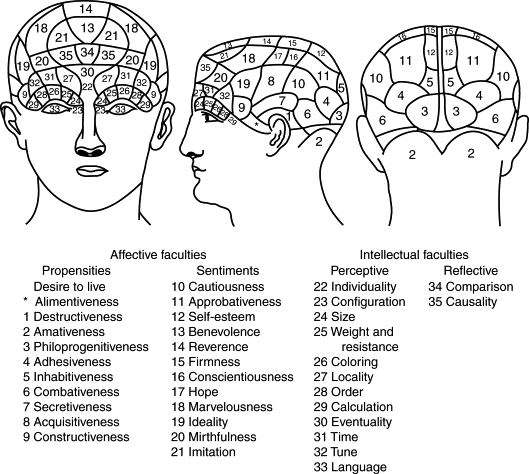
Franz Joseph Gall (1757-1828) achève la laïcisation du cerveau (rupture avec l’église) déjà bien entamée par La Mettrie et Cabanis. Il se distingue également des thèses sensualistes de John Locke (1632-1704) et Etienne de Condillac (1714-1780) (primauté des sens). En effet, véritable père de la phrénologie Gall va décrire 35 facultés morales et psychiques propre à l’être humain et les cartographier au cortex cérébral. Gall est un localisationniste mais ses méthodes sont très discutables, il va abandonner l’étude des circonvolutions ( dissection des aires du cerveau ) pour développer la science des crânes ou cranioscopie directement issue de la phrénologie.
Franz Joseph Gall
Etienne de Condillac
John Locke
Cartographie complète des fonctions du crâne par Gall (35)
Deuxième débat sujet à controverses: Unité ou fragmentation du cerveau ? Unitaristes contre Localisationnistes
Flourens (1772- 1844) dont les idées sont très appréciées par l’église et l’état va mettre Gall à rude épreuve. Il va pratiquer des ablations (méthode utilisée encore aujourd’hui) dont les résultats confirment les thèses localisationnistes mais va en faire une interprétation unitariste. Il considère que par l’ablation de certaines zones du cortex, toutes les fonctions générales disparaissent peu à peu (unité) alors qu’en fait chaque zone du cerveau à une fonction bien précise.
Pierre Flourens
En parallèle, François Leuret (1797-1851) et Pierre Gratiolet (1815-1865) vont établir les premières cartographies presque exacte des circonvolutions du cerveau. On leur doit la scissure de Rolando et la scissure de Sylvius et la découpe en différent lobes : pariétal, temporal, occipital, frontal.
Leuret
Gratiolet
Scissure de Rolando
Scissure de Sylvius
Jean baptiste Bouillaud (1797-1861) élève de Gall et continuateur de ses travaux va essayer de faire le lien entre langage et certaines aires du cerveau. Il invente l’anatomopathologie qui deviendra par la suite la neuropsychologie.
Jean Baptiste Bouillaud
Paul Broca (1824-1880), réussit cet objectif notamment avec le patient du nom de Leborgne dont on étudiera le cerveau après la mort. Leborgne n’arrivait pas à formuler une phrase mais répétait toujours « tan-tan ». Son cerveau présentait une lésion dans le lobe frontal de l’hémisphère gauche. On fait donc le lien entre la production du langage et cette zone corticale. Broca va donc cartographier toutes les zones du cerveau de manière fonctionnelle. Il va mettre en valeur une asymétrie entre les deux hémisphères du cerveau ce que ne soupçonnait pas Gall.
Paul Broca
Korbinian Brodmann (1868-1918) va ainsi proposer une carte fonctionnelle complète des aires du cerveau humain et du singe qu’on utilise toujours aujourd’hui.
Korbinian Brodmann
La carte des aires de Brodmann
Les localisationnistes s’imposent donc vers 1900 bien que certains unitaristes pensent toujours qu’ils ont raison. Ainsi il faut souligner le point de vue du docteur Head qui émet une critique plus nuancé du localisationnisme. Il explique que la localisation n’est pas intrinsèquement liée qu’au cortex et qu’elle est trop simpliste pour rendre compte intégralement du fonctionnement du système nerveux. Ses critiques vont stimuler encore plus la recherche des vainqueurs du débat.
Henry Head
Pendant ce temps, Van Leeuwenhoek (1632-1723) et Robert Hooke (1635-1703) découvrent la cellule dans les tissus végétaux et dans le sang. On cherche alors à répondre à la question suivante : de quoi la matière grise et blanche sont constituées ?
Van Leeuwenhoek
Robert Hooke
L’histologie du neurone
L’histologie est l’étude grâce aux microscopes, d’abord rudimentaires, au fil du temps les microscopes vont devenir de plus en plus précis en passant de l’optique à l’électronique.
Mais n’allons pas trop vite Malpighi (1628-1694) observe pour la première fois un cerveau avec un appareil grossissant en 1685.
Marcello Malpighi
Van Leeuwenhoek observe les nerfs avec un appareil d’optique rudimentaire si bien qu’ils croient que les nerfs sont creux. En fait il observe les axones composés d’une gaine de myéline (substance blanche).
Jusqu’en 1824 et Dutrochet (1776-1846), les observations au microscope ne progressent pas jusqu’à ce que que celui-ci découvre le corps cellulaire ou soma. Plus tard Valentin découvrira les queues protoplasmiques qui sont les dendrites. Deiters (1834-1863) dans une lettre posthume va conclure que le soma (voir aussi pericaryon) a un noyau et un cytoplasme que l’axone est unique et que les dendrites sont ramifiées.
Dutrochet
Deiters
Malheureusement ? Comment confirmer ? En effet les somas ne s’assemblent pas entre eux. Les neurones s’assemblent par les dendrites ce qui est impossible à voir au microscope optique à l’époque.
C’est ainsi que naît le troisième débat sujet à controverse :
Troisième débat sujet à controverses : Système nerveux continu ou contigu ? Réticularistes contre Neuronistes.
Joseph Von Gerlach (1820-1896) chef de file des réticularistes (voir la définition de réticule en médecine) identifie un plexus dendritique grâce à la coloration au chlorure d’or. En réalité ce plexus est un artefact et Golgi réticulariste tout comme lui qui enseigne l’histologie à Pavie met au point une coloration : la réazione néra qui révèle l’intégralité de la cellule nerveuse avec ses axones et ses dendrites. Spécialiste du cortex, il croit voir un réseau axonal continu et avouera en 1906 qu’il ne peut abandonner l’idée d’une thèse ou le système nerveux serait continu. En fait derrière les thèses réticularistes transpirent les thèses unitaristes.
Golgi
Joseph Von Gerlach
Réaction noire de Golgi
Première estocade portée à Golgi par Wilhelm Kuhne (1837-1900), en effet celui-ci décrit que quand le nerf moteur arrive au niveau de la fibre, il ne pénètre jamais dans le muscle ou cylindre contractile.
Wilhelm Kuhne
Wilhelm His (1831-1904), embryologiste suisse va porter un autre coup : en effet le neurone au stade embryonnaire est dépourvue de neurites qui sont les axones et les dendrites.
Wilhelm His
Viennent se mêler les psychiatres, Freud (1856-1939) de formation neurologiste fait des travaux d’anatomie qu’il pense venir au soutien des thèses réticularistes pendant que Forel (1848-1931) étudie la dégénérescence du réseau axonal et dendritique et analyse que les unités endommagés ne pénètre pas le corps cellulaire. (neuroniste)
Sigmund Freud
Forel
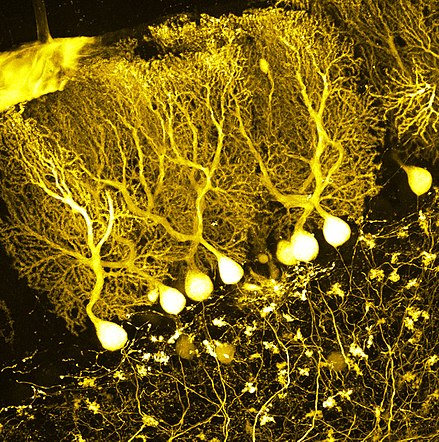
Le coup final à l’avantage des neuronistes est porté par Ramon y Cajal (1852-1934) grâce à un type de cellules spéciales qu’on appelle les cellules de Purkinje dans le cervelet qui possèdent un axone en corbeille du corps cellulaire et qu’il est anatomiquement indépendant de la cellule cible.
Ramon y Cajal
Cellules de Purkinje
La microscopie électronique confirmera les thèses des neuronistes dont le mot neurone et chromosome viennent de Waldeyer. Plus encore elle mettra en évidence le fait que le système nerveux est contigu et que l’espace entre ces neurones est comblé par la synapse dont le nom vient de Sherrington (1857-1952)
Waldeyer
L’électricité et les substances médicamenteuses
Francis Glisson (1577-1677) professeur de physique de l’université de Cambridge extrapolait déjà en réaction à la machinerie de Descartes, une thèse à propos de l’excitabilité des tissus nerveux ainsi qu’une sensibilité propre à ces nerfs.
Francis Glisson
Albrecht Von Haller (1708-1777) expérimentera à l’aide d’alcool, de potasse caustique et de vitriol le deuxième point : les muscles se contractent. Mais quelle est la substance qui provoque cette contraction ?
Von Haller
Luigi Galvani (1737-1798) qui travaille sur l’électricité atmosphérique utilise une barre de fer et implante la moelle épinière de la grenouille avec un crochet en cuivre et les pattes se contractent. Volta (1745-1827) son compatriote conteste, pour lui Galvani n’a prouvé que la conduction de l’électricité métallique et il a raison. On devra d’ailleurs à Volta la création de la pile et en son honneur l’unité de tension électrique.
Luigi Galvani
Alessandro Volta
Galvani réagit et enlève la moelle épinière du corps de la grenouille, dénude une cuisse et la met en contact avec la moelle, la cuisse se contracte encore. Voici une autre controverse entre électricité métallique et animale qui ne prendra fin qu’avec Mateucci en 1838 et grâce à un appareil électrique qui va enregistrer un courant propre issu du muscle : le galvanomètre dont le nom dérive de Galvani.
On va donc passer de la biologie à la physique et ce grâce à Du Bois Reymond (1818-1896) qui va porter la physiologie mécanique ancêtre de ce qu’on appelle aujourd’hui la biophysique. Il montre que le signal qui se propage le long du nerf est une onde de négativité qui se transforme en courant électrique et en potentiel d’action.
Du Bois Reymond
Von Helmholtz (1821-1894) applique son expertise de la balistique pour mesurer la vitesse du courant électrique et celui-ci est bien inférieur à la vitesse du courant dans un fil de cuivre et même du son (340 m/s) pour atteindre entre 25 et 40 m/s.
Von Helmholtz
Ainsi Mateucci et Flourens vont essayer de stimuler le cortex chimiquement et électriquement sur des grenouilles pour en activer les cuisses, mais rien ne se passe. Au grand plaisir de Flourens qui s’était déjà opposé à Gall et dont les thèses spiritualistes sont très en vogue à l’époque.
Mais Fritz et Hitzig deux médecins berlinois sont sur d’eux, ils ont vu un mouvement des yeux sur l’homme après stimulation corticale. Ils entreprennent alors des expériences sur les chiens, animal voisin de l’homme et constatent bien un mouvement oculaire. Mais comme dirait Volta, cela ne veut pas dire que le cortex génère sa propre électricité !
Caton (1842-1926) vient à leur secours, en 1875, il place une électrode sur la matière grise du lapin et avec son galvanomètre obtient une mesure du courant électrique. Lorsqu’il expose la rétine à une onde lumineuse il obtient un signal plus fort. Caton invente là l’électroencéphalographie(ECG) : Le cortex cérébral génère donc lui aussi des ondes électriques.
Richard Caton
Mais que se passe t-il alors entre la terminaison de l’axone moteur et le muscle ? Si les neuronistes ont raison que le système nerveux est contigu et non continu comment en rendre compte ?
La pharmacologie
Claude Bernard (1813-1878) en 1857 tente de montrer l’effet du curare utilisé par les amérindiens sur des flèches pour paralyser leur proie sur le système nerveux. Il postule la mort du nerf moteur une fois le curare introduit. Mais il se trompe, le curare ne fait qu’interrompre la communication entre le nerf moteur et les muscles. On suppose alors une substance chimique analogue au curare qui assurerait la communication dans le système nerveux.
Claude Bernard
Elliott apporte la solution avec la vessie du chat et sa stimulation par l’adrénaline isolée quelques années plus tôt. L’adrénaline se comporte donc comme un stimulant libéré chaque fois que l’influx nerveux arrive à la périphérie. Plusieurs dizaines d’années après, l’acétylcholine sera identifiée comme la substance produite par la cellule nerveuse qui assurent le transfert du signal électrique par la synapse.
Pharmacologistes et biochimistes vont alors découvrir dès 1940 par Martha Louise Vogt que de l’acétylcholine est présente dans le cortex central.
Martha Vogt
Von Euler (1905-1983) va isoler la noradrénaline que Hillarp (1916-1965) et ses collègues suédois vont détecter par fluorescence sur des coupes histologiques des neurones qui en contiennent. Le système nerveux se pense donc maintenant en cations sodium, potassium, calcium, molécules et en neurotransmetteurs.
Ulf Von Euler
Voilà donc l’histoire synthétisée des neurosciences d’Aristote jusqu’à peu de temps avant nos jours. Je m’attaquerai volontiers quand j’aurai plus de temps au fonctionnement général du cerveau de manière détaillée dans plusieurs articles à suivre au sein de la catégorie Neurosciences.
En guise de préambule pour notre catégorie sur les sciences cognitives, je souhaitais vous faire part d’une lecture très enrichissante sur la genèse de ces dernières.
Ecrite par un docteur en ingénierie informatique, également philosophe. Jean Gabriel Ganascia est un éminent scientifique qui a créé et dirigé le programme de recherches coordonné baptisé « Sciences cognitives » pour le compte du ministère de la recherche ainsi que le groupement d’intérêts entre ce ministère, le CEA, l’INRIA et le CNRS.
Rappelons dans un premier temps ce que sont les sciences cognitives
Le mot cognitif vient du latin cognoscere, qui signifie connaître. Les sciences cognitives décrivent donc tous les processus physiologiques et psychiques qui permettent à notre corps et notre esprit d’apprendre à connaître.
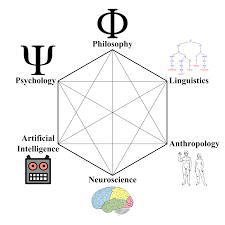
Ces sciences sont au carrefour de plusieurs sciences dures, sociales ou appliquées telles que les neurosciences, la linguistique, l’informatique, l’anthropologie et la psychologie.
@Wikipedia
Dans ce livre, l’auteur s’intéresse en premier lieu, aux origines des sciences cognitives loin dans le passé notamment avec les premières machines de Vaucanson qui illustraient certains processus physiques et physiologiques du vivant.
Puis les premières machines téléologiques (dont le but est une fin en soi) dites à rétro-action tels que le four ou le réfrigérateur furent créees. Le procédé de rétro-action est un principe issu de l’électronique, qui veut que la différence entre la valeur de sortie (température du frigo) et la température voulue en entrée entraine une régulation ou une action.
L’ancêtre officiel des sciences cognitives a pour nom la cybernétique et a été fondée par Norbert Wiener en 1947. Elle regroupe : simulation, rétroaction, information et a pour but l’étude des phénomènes physiques, sociaux et biologiques. De cette discipline on retiendra notamment la première modélisation des synapses et du neurone artificiel par Donald Hebb auxquels étaient affectés des coefficients numériques illustrant un mouvement : le connexionnisme.
Comparaison Neurone et Synapse de Hebb ici à droite
En parallèle, les premiers ordinateurs furent conçus et cela changea drastiquement la donne. En 1957 naquit au collège de Darmouth : l’IA ou l’intelligence artificielle dont l’approche va être d’assimiler le comportement psychique à un traitement de l’information.
Peu de temps après, les philosophes cognitivistes formulèrent un concept novateur : le fonctionnalisme calculatoire. Une analogie entre la machine et l’humain fut établie : le hardware étant notre cerveau, le software notre psychisme et les états de la machine nos états mentaux.
En 1980 avec l’avènement des neurosciences qui commencèrent à décrire les processus cérébraux par l’étude des neuromédiateurs qui agissent dans les liaisons synaptiques entres nos neurones, le neoconnexionnisme émergea et résolut beaucoup de problèmes.
Dans une deuxième partie le livre explique que les sciences cognitives sont au centre de trois pôles : psyché, cerveau et société. Il y décrit le concept de fonction qu’on retrouve dans beaucoup de sciences différentes et qui occupe une place centrale dans la cognition par exemple pour notre mémoire, nos sens, notre langage.
Je vous conseille de lire ce livre si vous vous intéressez de près ou de loin aux sciences cognitives.
Si vous n’avez pas peur de vous immerger dans plein de concepts, la finesse d’analyse de l’auteur vous permettra de mieux comprendre pourquoi les sciences cognitives sont fondamentales pour notre compréhension de l’humain dans le présent et dans le futur.
Pour ma part, j’ai toujours été fasciné par tous les mécanismes physiologiques et psychologiques qui nous permettent d’apprendre à mieux connaître, ou mieux se souvenir.