
Ceci est un résumé du chapitre 3 de Changeux « L’homme neuronal » :
Accrochez-vous c’est à mon sens un des chapitres les plus denses du livre l’homme neuronal de Jean Pierre Changeux en termes de concepts mais il est fondateur !
Electroencéphalographie
Après avoir étudié les différentes parties du cerveau, la manière dont elles se structurent entre elles de manière typologique et topologique sur un plan statique. Passons à l’étude dynamique de leurs fonctions : comment la machine cérébrale organise la transmission de signaux à travers les différents organes sensoriels et moteurs.
Nous savons notamment que le cerveau fonctionne grâce à des impulsions électriques, commençons par une étude d’électroencéphalographie :
Hans Berger 1873-1941

Hans Berger plaça pour la première fois des électrodes sur le cerveau humain contrairement à Caton et Matteucci qui avaient pris pour cobaye des animaux. Ces électrodes ne pouvant être implantés directement dans le cerveau, elles sont apposées à la surface du crâne, la traduction du signal étant optimisé par un amplificateur de gain.
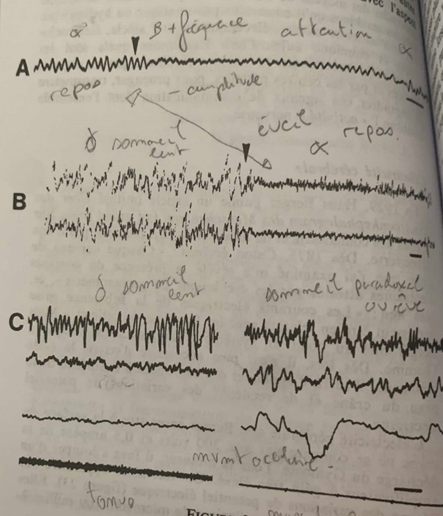
A l’état de repos la fréquence des ondes électriques est lente et de 10 cycles par secondes et leur amplitude est forte. Ce sont les ondes alpha α. Au contact d’un corps solide telle une tige de verre, la fréquence des ondes devient plus forte, la fréquence est plus forte et l’amplitude diminue : l’attention se fixe, ce sont les ondes Beta β. #ondes
Lors du sommeil lent l’amplitude des ondes est encore plus forte et la fréquence encore plus faible qu’à l’état de repos. Ce sont les ondes delta δ. Lors du réveil on passe des ondes δ à α donc à l’état de repos. #ondes
Un autre état est celui de sommeil paradoxal ou rêve, contrairement au sommeil lent les impulsions électriques sont souvent en rafales. Les yeux bougent et le tonus musculaire s’affaisse. #ondes
Cependant cette étude s’avère beaucoup trop généraliste de par l’irrégularité des ondes β et la similitude de fréquence et d’amplitude des ondes δ et α pour comprendre les phénomènes intrinsèques qui se produisent au niveau cellulaire et moléculaire. #ondes
On doit alors passer au cerveau de l’animal pour pénétrer le cortex avec l’électrode, on obtiendra en une fois ce qu’on a obtenu avec les électrodes apposées sur le crâne humain dont les résultats furent cumulés. #ondes
En utilisant une électrode dont la pointe est entre 0,5 et 5 micromètres plus petite que celle de la cellule nerveuse et on obtient des impulsions très brèves dont l’intensité s’accroît puis s’évanouit (discrètes) lorsque l’on déplace la microélectrode sur toute la surface corticale.
On répète l’expérience en plaçant l’électrode dans le cortex somatosensoriel, on stimule alors la patte du rat et l’on obtient une activité qui permet de mettre en lumière une source électrique forte continue et produite par des entités discontinues j’ai nommé le neurone.
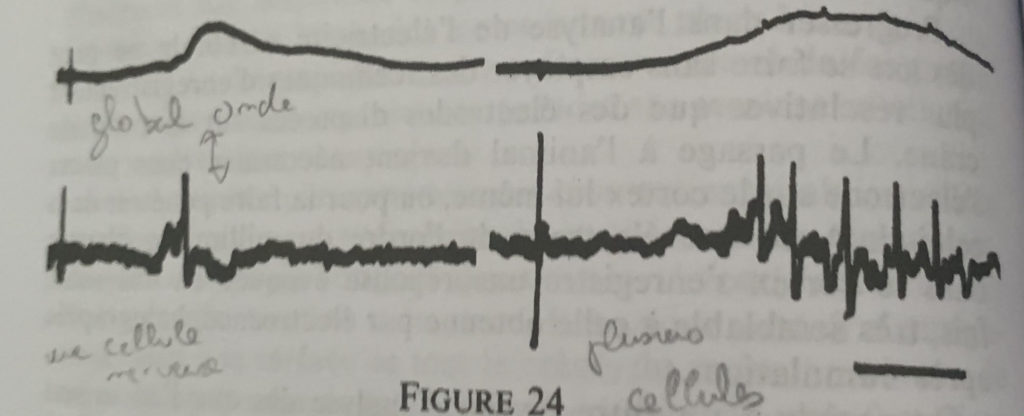
Cette démarche encéphalographique permet donc la mise en relation de l’activité électrique avec sa microstructure (neurone) mais ne pourra rendre compte du fonctionnement électrique du neurone de par sa multitude et des interactions complexes entre chacun.
L’influx nerveux
La vitesse de propagation électrique de l’influx nerveux chez les animaux primitifs se déplacent de 0,1 m/s (méduses) contre 100 m/s pour certains mammifères ce qui reste inférieur à la vitesse du son mais peuvent atteindre le mètre. La membrane cellulaire du neurone conduit le courant et remplace les fils de cuivre dans une communication électronique.
La durée de l’impulsion ne varie pas, une période réfractaire intervient pour maintenir sa durée ainsi que pour espacer deux influx successifs. L’amplitude des impulsions d’un dixième de Volt ne varie pas non plus. Quel que soit l’endroit où on l’enregistre ou la manière dont il est produit l’influx nerveux à la même forme.
La communication dans le système nerveux se traduit par un système nerveux très uniforme d’impulsions électriques voir universel.

Si l’on pénètre maintenant avec la microélectrode dans le neurone même on obtient un potentiel électrique stable dit de repos. La valeur varie par rapport à celle de la périphérie (Axone) de 0,50 à 0,90 mV.
Comment expliquer cette différence de potentiel avec l’expérience précédente à la périphérie au niveau de l’axone dans lequel le potentiel électrique était plus fort ?
Le neurone est une pile électrochimique
Le neurone est constitué d’atomes de sodium et de potassium, cependant on relève dix fois moins de sodium à l’intérieur qu’à l’extérieur mais dix fois plus de potassium. La chimie se transforme en électricité par ionisation, la libération d’un électron (particule charge négative) de valence va transformer le sodium et le potassium en ions positifs Na+ et K+. La membrane va filtrer ces ions et ne laisser passer à l’état de repos que les ions potassium de part et d’autre provoquant une force électromotrice négative à l’intérieur.
L’énergie qui maintient les concentrations d’ions potassium et sodium est une molécule biochimique : l’adénosine triphosphate ATP produite par respiration cellulaire (phénomène par lequel la mitochondrie (métabolite) transforme par phosphorylation (phosphore) oxydative l’Adénosine Di(2)phosphate en Adénosine Tri(3)phosphate). L’ATP Synthase qualifiée d’enzyme pompe de l’ATP (le suffixe ase désigne toutes les enzymes) sectionne ensuite l’ATP et cette scission va servir d’énergie pour provoquer l’influx nerveux de part et d’autre de la membrane.
Au repos la membrane est donc quand même mise sous tension par l’ATP mais elle ne laisse pas passer les ions sodium. La théorie du repos a été défendue par Julius Bernstein
Julius Bernstein 1839-1917

et confirmée plus tard par Charles Overton qui théorisera l’effondrement de la membrane par scission d’ATP pour expliquer la génération d’impulsion nerveuse.
Charles Overton 1865-1933

Huxley et Hodgkin vont préciser de manière universelle tout ce que nous venons de voir (mécanique ionique de l’influx nerveux) à partir d’une série d’expérience en utilisant encore l’axone géant du Calmar. En effet la perméabilisation de la membrane aux ions sodiums est commandée par une valeur seuil de potentiel électrique.
Les ions sodiums s’engouffrent dans des molécules canaux et le potentiel passe d’un signe négatif à un signe positif + 20 mv. L’impulsion électrique générée dépasse l’amplitude de 100 mV le long de l’axone. Ces canaux vont se refermer, d’autres canaux pour le transport d’ions potassium vont s’ouvrir et l’on retournera à l’état de repos, cette impulsion électrique aura duré 1ms.


Ceci s’applique universellement du nerf sciatique du rat à l’axone du Calmar et au cerveau humain.
Les oscillations
On a vu que même à l’état de sommeil lent au niveau encéphalographie, l’activité électrique était présente même sans stimulation sensorielle. Les neurones mis en cultures et tumoraux ont quand même un potentiel électrique régulier. Ces oscillations témoignent d’une activité spontanée du neurone.
Les expériences réalisées sur l’aplysie, la limace de mer qui présente l’avantage d’avoir des neurones géants vont faire état d’oscillations en rafales d’une parfaite régularité d’une dizaine d’impulsions toutes les 5 à 10 secondes.
Ilya Prigogine s’empare de la question pour voir si ces oscillations cadrent avec les lois de la thermodynamique son domaine scientifique de prédilection. Voici les trois lois établies avant lui par Lord Kelvin à laquelle obéit cette science macroscopique :
Ilya Prigogine 1917-2003

1/ L’énergie est égale dans l’univers, on ne peut ni créer ni détruire, on peut faire l’analogie avec Lavoisier qui dit que rien ne se perd mais tout se transforme. Ainsi on peut résumer cette loi par le syntagme d’Echange universel.
2/ L’Entropie augmente toujours dans un système isolé, l’entropie se caractérise par l’énergie thermique qui ne peut être utilisé d’aucune manière. Au sens général l’entropie est une grandeur propre au désordre de la matière d’un système.
3/ La troisième introduit le zéro absolu -276,15 C° et dit qu’à cet état l’entropie est nulle. Mais on sait qu’aucun être vivant rien ne peut vivre à cette température cependant on comprend que plus on tend vers une entropie faible, plus l’état énergétique est stable.
On parle même d’état métastable un peu comme la supraconduction qui expulse le champ magnétique et fait flotter le train hyperloop au-dessus des rails.
Comment les oscillations du neurone cadrent-t-elle avec ses trois lois ?
Je mettrai des chiffres à côté des mots qui vous permettent de relier ces conclusions aux 3 lois.
Première conclusion : les oscillations du neurone appartiennent à un système ouvert (2) dans lequel il y’a échange (1) constant (3) d’énergie. (De par la perméabilité de la membrane, dans lequel l’énergie s’échange grâce à l’ATP)
Deuxième conclusion : Les oscillations ne se développent jamais près de l’équilibre, pour cela il faut que le système soit hors d’équilibre*mais stable (3). Le neurone répond parfaitement à ces deux conditions (transfert inégale et continu d’ions)
Troisième conclusion : Des relations non linéaires* (propre des oscillations) doivent exister entre forces et flux, concrètement cela se présente lorsque des réactions se développent de manière explosive et lorsque des couplages s’établissent entre entrées et rétroaction (feedback en cybernétique) Le déclenchement de l’influx nerveux satisfait à ces conditions.
*Pour illustrer ces conclusions il est bon de souligner que Prigogine s’est écarté à la fin de sa vie de la thermodynamique classique a établi le concept de structure dissipative : « une nouvelle ‘structure’ est toujours la conséquence d’une instabilité. Elle est engendrée par une fluctuation. »
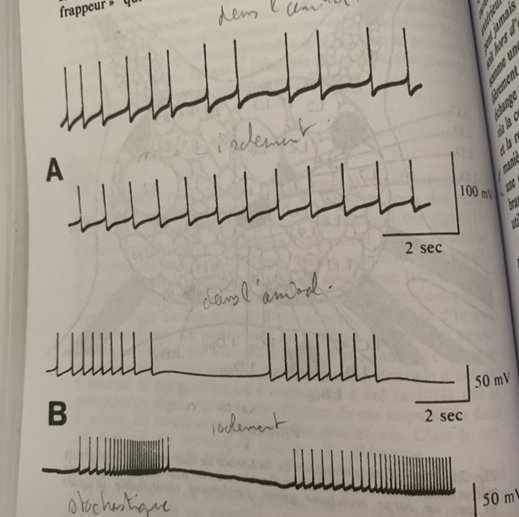
La particularité de cette oscillation est sa rafale :
Chaque rafale se greffe sur un générateur d’oscillation(pacemaker, rythme) dont le potentiel électrique oscille entre deux valeurs extrêmes de part et d’autre du seuil d’apparition de l’influx nerveux. Lorsque la valeur seuil du potentiel électrique est franchie une impulsion part et puis une autre tant que le potentiel reste au-dessus de la valeur seuil.
Comme pour l’influx nerveux on retrouve deux molécules canaux mais leur ouverture est de l’ordre d’une seconde pleine contre une milliseconde pour ce dernier. D’autre part le filtrage de la membrane du neurone est différent, les molécules-canaux filtrent les ions potassium et calcium (exclu de l’intérieur du neurone par une enzyme comme l’ion sodium) :
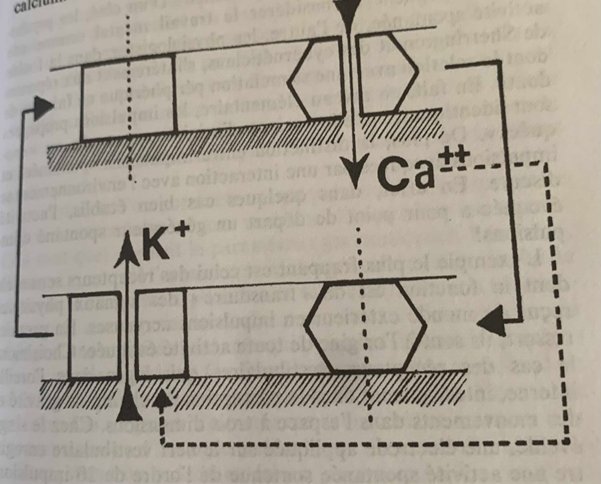
La canal calcium très lent s’ouvre par diminution du potentiel électrique crée par le canal potassium et laisse passer l’ion Ca++ qui une fois rejeté par l’enzyme pompe va provoquer l’ouverture du canal potassium dont on récupère l’ion K+ et on recommence comme dans un cycle. Ce couplage rétroactif est donc en parfait accord avec la troisième conclusion de Prigogine.
Si l’oscillation a une amplitude suffisante pour que la valeur seuil de l’influx nerveux soit franchie une rafale part à la crête de chaque oscillation lente : la longueur de la rafale dépend de l’amplitude et de la durée de l’oscillation.
Si le temps est bref on peut n’avoir qu’une impulsion voir pas du tout.
On a donc soit une horloge à rafale soit un générateur stochastique d’impulsion déterminé par la concentration de calcium dans la membrane.
Les sens et l’activité spontanée
Les récepteurs sensoriels ont pour but de transduire les signaux perçus par le monde extérieur en impulsion électrique : Lorsque l’on fait tourner un singe assis sur une chaise auquel on a placé une électrode dans le nerf vestibulaire de l’oreille dans un sens : on obtient 30 impulsions. Dans l’autre sens on obtient plus que 10 impulsions.
L’organe vestibulaire se compose de neurones tournés en direction du système nerveux central et de cils palpeurs qui baignent dans du liquide. Lorsque l’on tourne d’un côté la membrane va être orienté de telle façon que les canaux calciums s’ouvrent : diminution, ou se ferment : accroissement.
La variation d’un paramètre physique (gravitationnel, chimique ou lumineux) se trouve donc traduite en variations d’impulsions nerveuses. Les organes des sens se produisent comme des commutateurs d’horloge moléculaire. Les stimuli physiques les avancent, les mettent à l’heure où les retardent.
Les propriétés de générateurs d’impulsions ne sont pas réservées aux cellules sensorielles Il s’agit d’une propriété générale de la cellule nerveuse distribuée à de multiples niveaux.
La machine nerveuse contient donc de multiples générateurs d’impulsions distribuées tant au niveau du système nerveux lui-même que dans son ensemble.
D’un neurone à l’autre
Le sens de propagation des impulsions électriques est unidirectionnel, il part du corps cellulaire jusqu’à la terminaison de l’axone puis les dendrites comme dans un circuit électronique avec des sens obligatoires et des sens interdits.
On sait que les neurones et dendrites ne sont pas toujours en continuité les uns avec les autres. Certains neurones sont juxtaposés et séparés les uns aux autres. Comment peut-on expliquer la transmission du courant dans cette organisation discontinue ?
Comment expliquer cette polarité étant donné que lorsque l’on induit du courant électrique au milieu de l’axone le courant part dans les deux sens ?
Sherrington dont on a parlé dans l’histoire des neurosciences l’explique par la synapse en comparant un tronc nerveux ou le signal est bidirectionnel et l’arc réflexe unidirectionnel (stimuli sensorielle entraînant une réaction nerveuse comme un mouvement). Pour lui la synapse est une barrière intercellulaire : un nexus (espace proche) entre deux neurones.
Sherrington 1857-1952
Ceci va engendrer un débat entre les pharmacologues pour lesquels la transmission est chimique et les électro-physiologistes qui soutiennent la thèse électrique. Un compromis est trouvé : il existe des synapses électriques et des synapses chimiques.
Les synapses électriques sont symétriques et se placent entre deux neurones contigus de 2 nanomètres d’épaisseur, il y’a communication directe entre les neurones la plupart du temps sans polarité de manière multidirectionnelle comme une horloge qui mettrait les neurones à la même heure.
La transmission chimique est paradoxalement découverte par un électro-physiologiste dont on a déjà parlé dans l’article premier Du Bois Reymond mais aussi aux pharmacologues Elliott, Sir Henry Dale, et Langley qui prélèvent sur certains organes cibles de la grenouille un ester de la choline, l’acétylcholine.


Ce neurotransmetteur a été découvert dans la nature à partir de l’ergot de seigle et synthétisé par Crum-Brown et Frazer


Faisons un petit exercice de biochimie en considérant la formule brute, la formule semi-développée et la formule développée de l’acétylcholine en partant de la composition d’un ester et de la choline.
Formule Brute Acétylcholine : C7,H16,N+O2
On trouve donc 7 atomes de carbone, 16 atomes d’hydrogènes, un atome d’azote, et deux atomes d’oxygène.
Voici la formule semi-développée qui présente une première disposition dans l’espace de manière rectiligne ainsi que les liaisons entre les atomes.
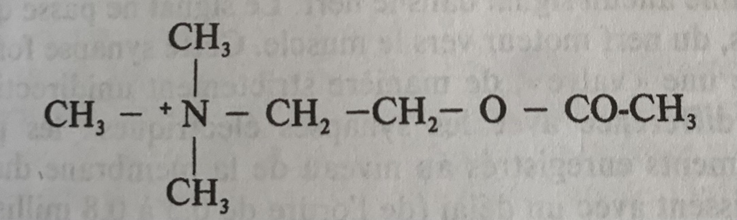
Comparons avec la choline formule brute C5,H14,N+O :
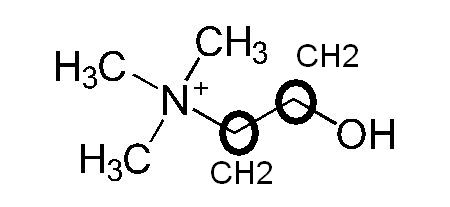
J’ai complété la formule développée en deux dimensions que l’on trouve sur wikipedia à partir de l’énoncé de la formule brute avec des cercles et des liaisons atomiques.
on retrouve les liaisons CH3 autour de l’atome d’azote, les deux sommets cerclés n’étaient pas notés atomiquement, je les ai rajouté.
En chimie, la fonction ester désigne un groupement caractéristique formé d’un atome lié simultanément à un atome d’oxygène par une double liaison et à un groupement alkoxy
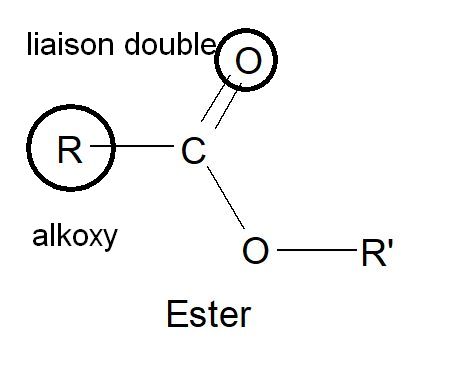
Lorsque l’atome lié est un atome de carbone , on parle d’ester carboxylique, dont la forme générale est R-COO-R’.
En chimie la lettre R est capitale, elle signifie Radical et sert parfois à désigner un groupement d’atomes mais aussi parfois dans le cas d’une chaîne latérale de fonctionnaliser une molécule.
Cette fonctionnalisation sert à caractériser un groupe d’atome qui chaque fois présent dans une structure moléculaire différente en modifie les propriétés chimiques de manière analogue.
Elle permet également de réprésenter les chaînes carbonnées en chimie organique. Le methyl a un atome de carbone quand l’éthyl en a deux.
Comparons maintenant la partie ester de la formule moléculaire de l’acétylcholine brute avec sa formule développée en 2D sur Wikipédia : C7,H16,NO2
on retrouve la double liaison caractéristique entre l’atome d’oxygène et l’atome de carbone, l’ester d’acétate se caractérise par la chaîne O–CO–CH3 :
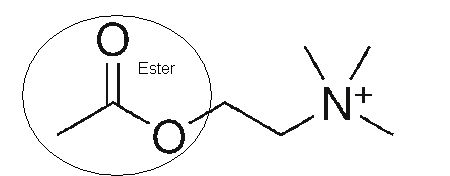
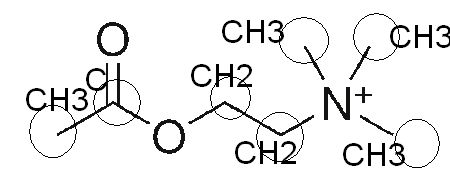
Reprenons ou nous en étions :
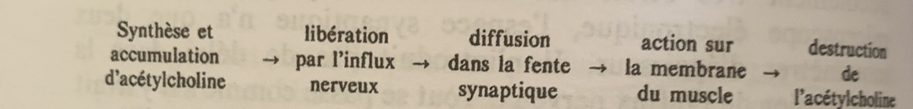
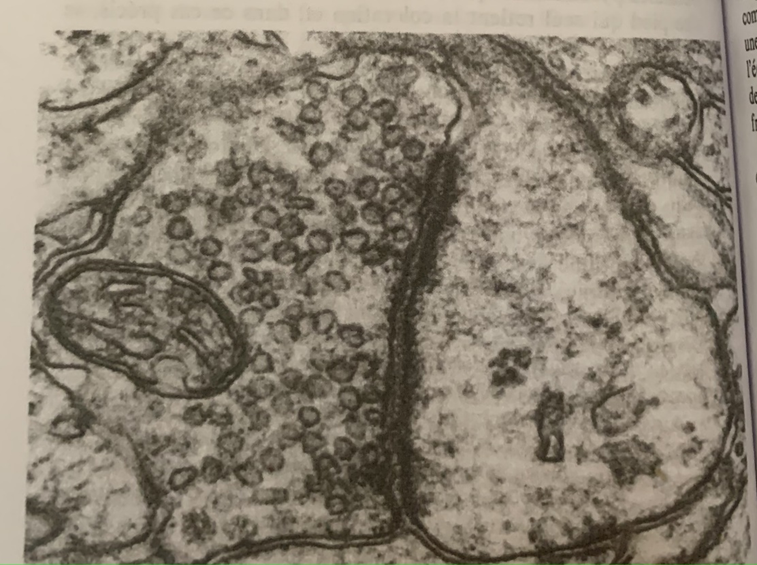
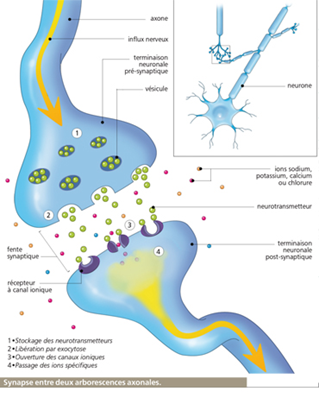
Cette molécule est libérée par l’influx nerveux entre le nerf moteur et le muscle strié. A la différence de la synapse électrique, la synapse chimique est polarisée (directionnel) et l’espace entre le nerf moteur et le muscle strié est 10 à 25 (20 à 50 nm) fois plus grand que l’espace entre deux neurones électrique.
D’autre part, leur morphologie est différente, la synapse chimique est asymétrique : d’un côté une terminaison nerveuse bourrée de vésicules de 30 à 60 nm pleines du neurotransmetteur. De l’autre un épaississement de la membrane ou densité post-synaptique.
Ces synapses révèlent la présence d’électricité générée par les impulsions issues de l’influx nerveux qu’elle transmet à la fibre musculaire de manière unidirectionnelle. Ces synapses fonctionnent comme une valve. Le signal ne fonctionne que dans un sens du nerf moteur au muscle.
On observe un délai entre 0,3 et 0,8 ms à la mesure de l’électricité dans le muscle, plus long que celui de l’influx nerveux : relation entre vitesse, distance et temps puisque le neurotransmetteur n’est pas présent dans le muscle à la base ce qui explique la polarisation.
Mécanisme de libération de l’acétylcholine
A la terminaison nerveuse, au niveau de la jonction nerf-muscle on sait que l’impulsion électrique provoquée par l’influx nerveux provoque la libération de 300 paquets d’environ 10.000 molécules chacune d’acétylcholine ce qui représente 3.000.000 de molécules.
Ce nombre est en définitive petit comparé à l’unité de matière basé sur la constante d’Avogadro qui est de 6,02 x 10^23 molécules dans une mole de matière mais dans un volume si petit que cette concentration est relativement forte.
Au repos, on a donc 10^-9 mole par litre d’acétylcholine mais lors du passage de l’impulsion nerveuse cette valeur est donc comprise entre 10^-4 et 10^-3 moles par litre pour une durée de l’ordre de la milliseconde puis s’évanouit grâce à une enzyme spécialisée : la cholinestérase.
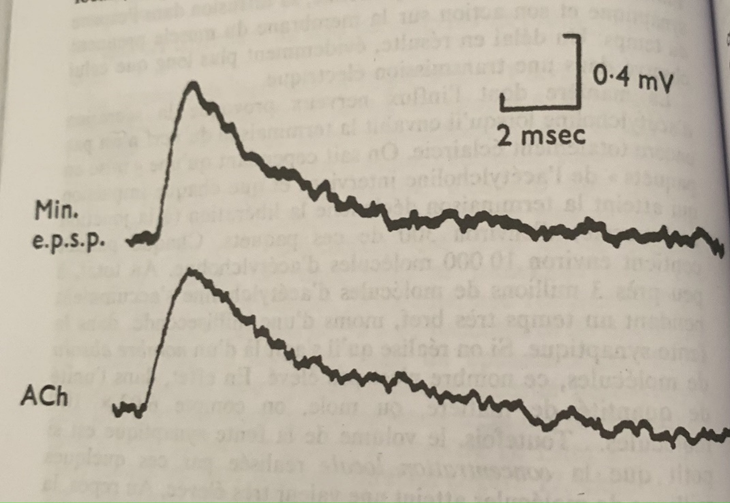
Retour d’une onde électrique
De l’autre côté de la terminaison nerveuse, au niveau de la face post-synaptique on enregistre une onde électrique bien différente de l’électricité générée à la base par l’influx nerveux. Son amplitude est 5 à 10 fois plus faible et sa durée est à 3 à 5 fois plus longue. (3 à 5 ms).
Cette onde résulte de la perméabilisation de la membrane de la synapse qui engage les ions sodiums, potassiums et calciques individuellement et non séquentiellement. (Un canal s’ouvre individuellement sans lien commun avec les autres et brutalement (1ms) (le caractère individuel rend plus simple la transmission du signal)
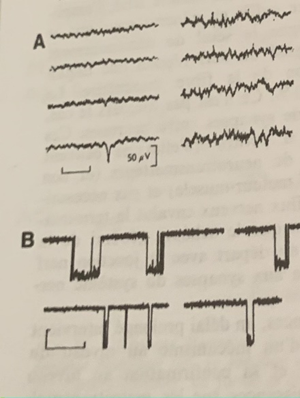
En B l’ouverture et la fermeture brutale unique (carré) des canaux sodiums, potassiums, calciums bien individualisés résultant de l’onde électrique générée par la transmission d’acétylcholine.
Résumé synthétique de la multiplicité des signaux
La libération de l’acétylcholine par l’influx nerveux entraîne la transformation d’un signal électrique (1) en signal chimique (2) L’ouverture des canaux ioniques constitue l’étape inverse et la transformation du signal chimique (3) en signal électrique (4) que j’appellerai onde de retour : elle résulte de l’ouverture de canaux au niveau de la synapse et est réencodée vers le premier neurone de la terminaison neurale pré-synaptique par lequel l’influx nerveux est passé afin de lui communiquer que le signal a été transmis.
Synthèse morphologique
Au niveau de la jonction nerf muscle strié, le potentiel déclenché par l’influx nerveux et la réponse synaptique dont résulte la transmission d’acétylcholine suffit à déclencher un influx. A chaque impulsion du nerf répond une contraction du muscle strié. La transmission est efficace à 100 %. Ce n’est pas toujours le cas en revanche pour des synapses entre deux neurones.
Ces terminaisons sont si petites qu’elles ne peuvent libérer qu’un seul paquet d’acétylcholine et pas nécessairement à chaque fois qu’un influx nerveux envahit la terminaison. Il n’en reste pas moins que le schéma général de la transmission chimique établi au départ avec la jonction nerf moteur muscle strié s’applique au système nerveux central.
Types de neurotransmetteurs
Franck Campbell McIntosh 1909-1992

L’acétylcholine a été isolé dans le cerveau pour la première fois par McIntosh en 1941 et la noradrénaline par Vogt en 1954 et ces deux neurotransmetteurs sont les membres d’honneurs d’une liste de substances chimiques qui s’allongent de jour en jour tels que :
- Acides aminés
- Amines biogène « double fonction » : dopamines, sérotonine…
- Polypeptides (Enképhaline, Endorphine, Substance P…
- Hormones : Somatostatine, Endorphine
- VIP (Peptide Vasoactif Intestinal)
Ce dernier a été découvert dans l’intestin ce qui montre bien que l’interaction entre le système nerveux périphérique et le système nerveux central.
Un autre exemple est la somatostatine qui bloque l’hormone de croissance et qu’on retrouve dans le pancréas.
On retrouve ces neurotransmetteurs en particulier chez tous les vertébrés. Aucun neurotransmetteur n’est propre à l’être humain. (Chimie prégnante dans le vivant)
La diversité biochimique qui ne cesse de s’accroître au fur et à mesure des recherches en neurosciences contraste avec l’uniformité du signal électrique. Un même neurone peut recevoir des dizaines de milliers de synapses et que ces synapses peuvent employer des neurotransmetteurs différents.
Cette diversité dans la nature chimique et dans les effets ioniques introduit une combinatoire de signaux qu’un neurone électrique n’effectuerait pas de la même manière ou n’effectuerait même pas du tout.
Les molécules serrures
La synapse chimique tient une place centrale dans la communication entre deux neurones, Elle canalise le transfert de signaux de cellule à cellule en créant une polarité dans ce transfert, crée des circuits et introduit une diversité au niveau de la membrane du neurone. Ses dimensions sont de l’ordre de grandeur d’une cellule bactérienne.
Afin de démonter le fonctionnement de la synapse chimique, il convient d’étudier l’effet du neurotransmetteur sur la face post-synaptique. Hors Emil Fischer postule en 1906 que pour qu’une molécule puisse agir sur un corps, elle doit pouvoir se fixer.
« Corpora Non Agunt nisi fixata »
Emil Fischer 1852-1919

Comment cette fixation se fait-elle ?
La cellule contient une substance chimiquement active (un récepteur) qui porte une configuration géométrique adapté complémentaire de celle du corps considéré (le neurotransmetteur). La métaphore est celle de la clé dans une serrure.
Langley montre que cette substance répond au curare (effet inhibiteur) à l’acétylcholine et à la nicotine de manière analogue et qu’elle n’est localisée sur le muscle adulte qu’au niveau de la terminaison nerveuse : c’est la serrure ou la molécule réceptrice.
John Newport Langley 1852-1925

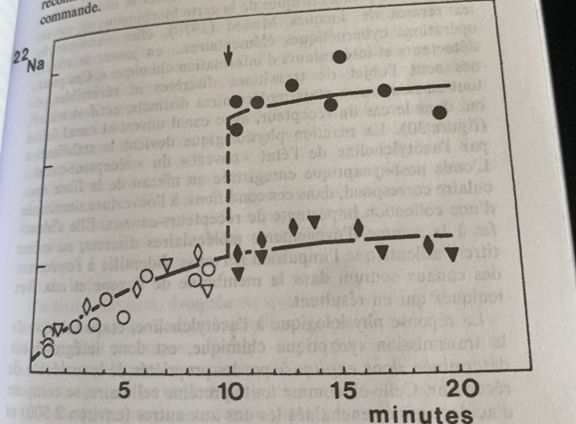
Comment le prouver ?
La nature recèle de richesse et le poisson « torpille » aussi appelé « gymnote » dont trois décharges de 500 V à 0,5 A tuent un homme résultent de la mise à feu d’un milliard de synapses très semblables par leurs propriétés à celles que l’on trouve à la jonction nerf-muscle de l’humain. L’organe électrique se compare à un muscle dont l’appareil contractile aurait été enlevé pour laisser les synapses à vif.
Cette accumulation gigantesque va se révéler fort utile pour le biochimiste. En plus ces synapses ont toute la même composition chimique : travailler sur 1 kg d’organe chimique revient à travailler sur une synapse géante qui pèserait le même poids.

Dans un premier temps on va utiliser comme produit radioactif pour marquer la face post-synaptique et détecter la forme et la position de cette fameuse molécule réceptrice le neurotransmetteur lui-même (acétylcholine). L’expérience échoue car de nombreuses molécules lie de manière non sélective à beaucoup trop de molécules du récepteur lui-même.
La nature va encore venir au secours des biochimistes : les serpents cobra et bungare ont une mauvaise réputation de par la toxicité de leur venin qui paralyse les voies respiratoires au même titre que le curare des Amérindiens. Cette toxine alpha qui bloque les récepteurs va être utilisé comme marqueur radioactif pour détecter la localisation sur de la molécule en question.

On trouve des serpents marins mangeurs de poissons torpilles dans la mer du Japon, leur réunion dans le tube à essai va permette d’isoler cette fameuse molécule réceptrice sur laquelle se fixe l’acétylcholine.
Cette molécule isolée a une masse de 250.000 soit 3,5 fois plus que celle de l’hémoglobine, elle se compose de plusieurs chaînes de quatre types différents dont une répétée deux fois. Elle est disposé sur toute la membrane sous-synaptique de la face post-synaptique.
Leur forme géométrique étudiée par centrifugation de broyats d’organes électriques est celle de micro sacs que l’on peut remplir d’ions sodiums ou potassiums radioactifs pour mieux comprendre l’effet de l’acétylcholine qui provoque l’ouverture des canaux.
Ou se trouvent ces canaux ioniques ? On disperse les micro-sacs grâce à des détergents sans les détruire dans le tube à essai. Elle garde ainsi toutes ses fonctions natives comme sur la membrane sous synaptique. On peut ainsi en étudier son électrophysiologie qui renvoie plusieurs signaux exactement similaires à l’ouverture de plusieurs canaux individualisés brutalement (1 ms) de forme carré.
La molécule de masse 250.000 contient à la fois le site récepteur et le canal ionique.
Cette molécule est analogue à une protéine allostérique, ce concept défini par Jacques Monod et Jean-Pierre Changeux veut que la fixation d’une molécule effectrice sur un site modifie les conditions de fixations d’une autre molécule en un autre site distant de la même protéine un peu comme deux particules quantiques intriquées.
Ces protéines sont sujettes à des transitions discrètes entre états moléculaires distincts actifs ou inactifs. Dans notre cas, le récepteur, l’illustration est l’ouverture et la fermeture ultra-rapide des canaux. #carré
L’onde post-synaptique enregistrée au niveau de la fibre musculaire correspond donc à l’ouverture simultanée d’une collection importante de récepteurs-canaux. #fibre
Elle s’identifie à la somme d’évènements moléculaires discrets au même titre que l’impulsion nerveuse s’identifie à l’ouverture de canaux sodiums dans la membrane de l’axone et aux flux ioniques qui en résultent.
La molécule réceptrice comme toute protéine cellulaire se compose d’acides aminés enchaînés les uns aux autres (environ 2500) Chaque acide aminé contient entre 10 et 30 atomes qui déterminent intégralement sa réactivité chimique, ses possibilités de liaison avec ses congénères (polymérisation) (plusieurs parties réunies) pour construire son édifice protéique (macromolécule) (Ex : peptide)
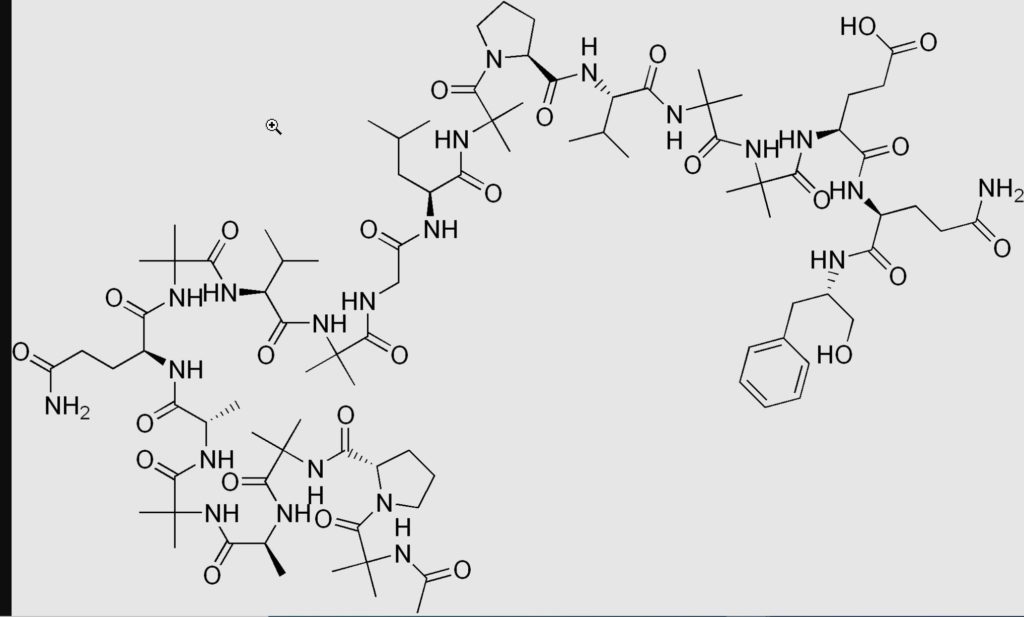
Comparez la structure moléculaire d’un seul acide aminé avec celle d’un peptide et vous comprendrez la polymérisation et la constitution d’une protéine que l’on peut qualifier de manière réductrice de macromolécule.
Conclusion transitionnelle
Nous accédons donc à une organisation au niveau moléculaire, atomiques et ioniques de la transmission chimique entre deux neurones.
Synthèse
Les atomes psychiques réexaminées (Démocrite)
La même méthode en définitive, conduit de l’enregistrement de l’encéphalogramme à celui des impulsions électriques produites par des neurones uniques, de la réponse post-synaptique aux transitions d’ouvertures des canaux ioniques, eux même identifiables à des changements de structure moléculaire.
A chaque étape il y’a réduction d’un niveau d’organisation à une unité plus élémentaire : de la population des neurones corticaux, à la cellule ou à la synapse, de l’impulsion nerveuse (ou à des courants post-synaptiques) à la molécule.
L’étude dynamique des différentes pièces du cerveau sont donc descriptibles avec les mêmes termes que ceux du physicien et du chimiste.
On pourrait plus loin et décrire la formule atomique de la molécule réceptrice dont on vient d’étudier le fonctionnement pour comprendre sur quoi se base sa réaction à l’acétylcholine.
L’activité spontanée et évoquée est donc directement issue de propriétés atomiques. Doit-on alors réexaminer les atomes psychiques de Démocrite ? Les ions sodiums et potassiums qui traversent les canaux de l’axone sont les mêmes que ceux que l’on trouve dans l’eau de mer ou à l’intérieur du neurone.
Les molécules de neurotransmetteurs sont composées de carbone, d’hydrogène, d’oxygène et d’azote qui n’ont rien de propre aux êtres vivants. Le système nerveux se compose de et emploie pour fonctionner – la même matière que le microcosme.
Celles-ci s’organise en édifices moléculaires qui interviennent dans la communication nerveuse au même titre que d’autres règlent la respiration cellulaire ou la réplication des chromosomes. Les protéines tiennent la une place critique puisqu’enzyme pompes (destruction ou aspiration), enzymes de synthèse des neurotransmetteurs et leurs récepteurs sont des protéines.
Au lieu de reprendre le terme d’atomes psychiques, doit- on parler de molécules psychiques ? Le trait le plus frappant qui se dégage des recherches actuelles, c’est que l’électricité et la chimie du cerveau sont les mêmes que celles des organes périphériques. On retrouve dans la nature : ce qui est vrai pour l’organe électrique du Gymnote est aussi vrai pour Homo Sapiens.
Au niveau des mécanismes élémentaires de la communication nerveuse, rien ne sépare l’homme des animaux. Aucun récepteur, canal ionique ou neurotransmetteurs n’est propre à l’homme.